|
|
3 STReTCH ModuleContents 3.1 Introduction - Statement of objectivesDespite a wide range of development strategies - architectural models sensu (Hallé, Oldeman et al. 1978) - all trees face the same fundamental constraints in terms of light capture and notably need to strike a balance between investment in support structure and assimilatory organs. The objective of the Stretch module is to propose a generic model to represent crown shape flexibility in response to light and space limitations independent of the detailed tree architecture. We believe that a proper crown model should in particular be able to allow for the simulation of a number of “typical” development trajectories. Responses expected to be covered by the new crown model include:
In the Stretch approach the crown is represented by a growing deformable solid. This expanding polyhedron is defined by a set of vectors (later referred to as “Virtual Branches”) all stemming from the crown base. The growth rate of those Virtual Branches (VB) is a function of (local) light conditions (local response) and their relative position (to capture the crown elongation a species specific characteristic). The way in which VBTs are affected by (local) light conditions or constrained by their relative position within the crown is species dependent. 3.2 Ecological basis – Biological PrinciplesWhile overall growth - as captured by the dbh increment – will decrease under sub-optimal or supra optimal light levels, crown shape may also be affected by sub-optimal or anisotropic light and in return affect overall growth performance. Two major mechanisms may contribute to crown shape alteration under specific light conditions A/ Local asymmetric competition between branches within a tree The so-called branch autonomy theory states that the local carbon balance between production and demand for growth and respiration determines the fate of the branch and notably whether it will be shed. However there is increasing evidence that such a simple view is not tenable (Henriksson 2001; Sprugel, Brooks et al. 2002; Lacointe, Deleens et al. 2004). Notably it was shown that the light level at which a branch will be shed depends on the relative light level (to the rest of the crown) rather than the absolute level of light experienced. Even though dominant trees have more resources to allocate, branches on suppressed trees are able to grow and produce new foliage at solar irradiances where branches on dominant trees die. Thus branches are sufficiently interdependent that a positive carbon budget by itself does not ensure branch survival; branch position relative to other branches on the same tree is also important (Sprugel et all, 2002). Furthermore, the increased growth of non-shaded branches in trees where only two branches were shaded suggests that resources were preferentially allocated to branches in more favorable positions (Henriksson 2001). Hence we expect that local deformation of a crown (for example the opportunistic development of a part of the crown in response to local abundance of light (side gap, row planting etc) will be better modeled as a combination of the overall growth potential modulated by local gradients. B/ A whole tree active shade avoidance response under low light/strong vertical light gradient by which crown growth is reoriented towards height at the expense of lateral growth and which is commonly observed under high tree population density or low light levels. This response may result from a combination of biological mechanisms. Relative or absolute increase of growth in height may also be a response of internal competition of allocation of “growth potential” within the crown as a decreasing gradient of light (or space!) availability from apex to base is common. Hence the distinction made here between global or local response is somewhat arbitrary and is made for operational purposes. Vertical light gradient of increasing intensity towards the top of the canopy is not only common under dense planting where overhead light is abundant while lateral light is very much restricted but also in the forest understory where a similar gradient (though probably less pronounced) may prevail in many cases. Hence new leaves are produced where light resource is most abundant (a local response) which translates into a global deformation of the crown: only the upper-most part of the crown receives adequate light to maintain active growth and therefore elongation of crown occurs. The strategy of "compression" in the understorey and accelerated growth under gap as described in (Sterck 1999) for example would indicate that for canopy species whose juveniles start to grow in the understorey, the relevant signal to trigger accelerated growth in height might be the light gradient rather than the light level… It has been shown that poplar trees can alter their growth rates under modified red/far red ratio and noticeably increase their relative growth in height (Gilbert, Seavers et al. 1995). This may be a widespread response to shading (Ritchie 1997) however our observations do not clearly support the assumption that the overall light level (which is correlated to the red-far red ratio) is associated with enhanced growth in height and this is not implemented in the present model. 3.3 Crown shape modeling3.3.1 analytical frameworkCrown development is decomposed into vertical extension and horizontal extension of VBs (“Virtual Branches”), and light may affect each directional component differently. Growth of an individual VB will depend on
3.3.2 The Algorithm3.3.2.1 Vertical stretching of crownCompute vertical and horizontal growth component of VBs of reference tree based on species specific shape and actual overall growth reducers Computation of stem height and dbh increments are described See section 2.3 Height Increment. Those values are then used to compute crown stretch by computing VB increments based on crown profile 3.3.2.2 A/ half ellipsoid profile(vi_a) VB_incr_ver=cos (theta) * height_inc (vii_a) VB_inc_hor= sin(theta) * a* dbh_inc where a refers to the linear relation between dbh and crown width i.e. crown_width = a*dbh + b and theta is the angle of VB with vertical If VB_Tip is inside neighboring crown horizontal component is set to 0 (but height increment is still applied to ensure decent crown profile). 3.3.2.3 B/ Conical profileAssumptions identical to above (implicit assumption: dbh
=dcbh)
let L’=L + height increment and H’= expected crown radius at crown base for new
and the current increment in length of VB of angle q is computed as l’-l 3.3.2.4 Step 2: lateral deformation of crownBased on the sky map and the light model adjust VB_inc_hor only for anisotropy of incoming light. At this stage we assume that the number of VBs per tree is fixed and set to the following:
Then the default number of azimuths of the sectoral light map is set to 15 (same as the number of VBs for a given inclination. The number of inclinations of the sectoral light map is set to 5 (equal to number of inclination in the light model, medium precision). Orientations of VBs is randomized by choosing the first VB randomly. Number of VBs and sectoral light resolution is not accessible to user
For each of the (15-1) * 15 VB directions (excluding vertical VBt) we compute
a index of efficient lighting for each direction and the average value of the
index. Note that this is done for all directions (whether there is or not a corresponding
VB alive). The ratio of this light level to the average light level is used
to adjust the horizontal component of growth for each growing VB. Note that
whether there are VBs missing or halted does not affect the deformation of the
remaining VBs. Let G(i) be the standard growth rate (equal for all azimuth) computed in
previous step Step 1 Step2 Finally if VB is inside neighboring crown (i.e. has intruded a neighbouring crown G’(i) is set to zero (no growth through neighbor's crown envelope) 3.3.2.5 Step 3: branch sheddingBranch shedding always starts with lower most VB in any azimuth. 1/ A first test for VB survival is made prior to VB growth. If VB sectoral
light-level
is below a certain threshold then it is dropped. Threshold light level at
which lower most VBs are shed should be dependent on overall light level (i.e.
be relative
to overall light level) and more responsive crowns (more flexible, more sensitive)
should drop their lower most VBs more rapidly. This is partly covered through
step 2 but only for well developed crowns (crown which are severely constrained
Note that the above formulation does not link branch shedding to reallocation of growth pattern but only to the perceived light gradient (via sensi) and thus is not entirely satisfactory. However sensitivity and flexibility should be correlated (highly sensitive non flexible is a combination that does not make sense). Further more the relative decrease in growth rate is somewhat arbitrarily computed (and trimmed). Sensitive species should deploy a shade avoidance strategy and should correlatively likely have higher minilum, higher flexibility, and (?) lower porosity. In the present form high flexi values (high crown plasticity) seems to confer an advantage in terms of rapid regrowth of crown (compensation growth) as flexibility will likely allow for more than proportional crown surface increment and faster recovery if a gap happens to occur in the vicinity of an otherwise strongly constrained tree for example (provided it is coupled with a high sensitivity to detect the gap…) 2/ Once VB have grown the crown surface is checked against
the expected
crown surface (cf allometric relation between dbh and crown volume in open grown NOTES:
3.4 Collision detectionCollision determines halt of growth of VB The collision between neighboring crowns is detected if there is intersection between the horizontal vector joining VBT to tree crown vertical axis and any triangle defined as a result of triangulation of VBTs location in 3D of neighboring tree. Note that as a result of this implementation vertical growth rate of tree top apex is not affected by collision. However the VB growing from inside another crown will be halted if they come to intersect with containing crown envelope. 3.5 Triangulation algorithm A proprietary triangulation algorithm is used which takes advantage
of the fact that VBs are regularly spread and notably that on a given
azimuth
there
can be Let n be the number of azimuth and p the number of inclinations. The total number of VBs (including apex) in a full crown (no missing VB) is then n*(p-1) + 1 and the associated number of triangles is n*(2(p-2) + 1)=n* (2p-3). Each time a non-vertical VB is dropped, so are two triangles as can
be seen from the algorithm below and illustrated in the figures.
The algorithm:
Let H(i,j) be a VBTs height at inclination index i and azimuth index
j. (assuming that each VBTs has height and horizontal distance from
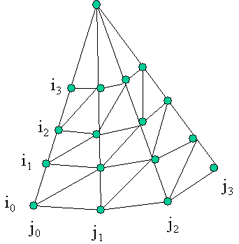
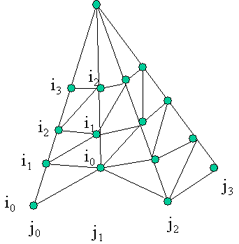
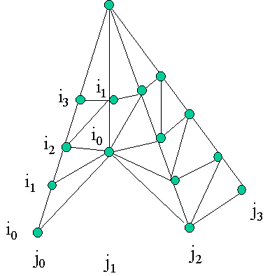
axis) The following algorithm iterate the VBTs through the inclination index on the two following series of VBTs per azimuth directions H(Ia, Jc) and H(Ib, Jc+1). Step a: Find next element Min(H(i1, Jc), H(i2, Jc+1)), store it (VBTs with the same index) as the second element of triangle. Step b: Else increase the i of the lowest VBT series and find next Min(H(i1, Jc), H(i2, Jc+1)) as the third element of triangle. Store the last two elements of the previous triangle as the elements for the next triangle. If there is more than 1 VBT left in both series then Go to step b. Else put the last VBT as the third element, and Figure 1. left figure shows the initial VBTs connection (subpart of a crown VBTs), and the right figure show the possible VBTs connection after a VBT shed at i=0 and j=1. the series of VBTs at j1 then re-indexed for i.
Figure 2: if 2 VBTs are shed.
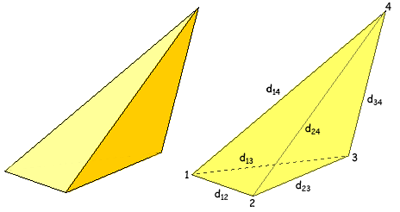
3.6 Algorithm for crown volume computationVolume of the crown is calculated as the sum of all connected tetrahedra, one face of which is a triangle of the crown envelope (as a result of the triangulation described above). Each triangle is then connected to the center of crown base (added to each triangle as one other vertex to form a tetrahedron). Then tetrahedron volume is calculated using the formula below: http://mathforum.org/dr.math/faq/formulas/faq.irreg.tetrahedron.html
3.7 Crown deformation and the pipe model theoryWe may need to explore further the application of the pipe model in order to link more formally crown volume and dbh values. To maintain consistency between overall crown volume and tree diameter we further assume that leaf area and stem cross sectional area are linearly related. This allometric relationship based on the functional relation between sapwood area and leaf area, is expected to be robust (Morataya, Galloway et al. 1999) and hold under the following provisions
Then LA is further broken down into crown volume and leaf area density (per unit volume). Assuming LAD to remain stable across time and space within a particular tree this implies that we can extend the allometric relation between stem diameter and LA to stem diameter below crown and crown volume (or crown surface if we consider that leaves are predominantly located on a thin layer on the outer most side of the crown). This relation is used to enforce branch shedding under extreme deformation (elongation) of tree in response to light gradient, i.e. as crown volume is constrained the lower most branches are shed. A tapering equation could be used to link dbh and diameter below crown so that the assumption of linear relation between stem cross sectional area and leaf area would be more robust. In first approximation a conical truncated shape may be used (based on data collected by Hubert in Krui for example) for the part between diameter at breast height and diameter at crown base height (but see also, for a discussion of the various approaches that may be used, http://sres.anu.edu.au/associated/mensuration/shape.htm#equation). In fact the model should be able to compute cross-sectional area of stem at any height based on any kind of stem profile if such information is provided by user |
||||||||||||||||||||||||||
|
Update 10-06-2005 Comments and questions send to: |
© Copyright ICRAF 2002 -
2005 http://www.worldagroforestrycentre.org/sea |