An e-publication by the World Agroforestry Centre
AGROFORESTRY A DECADE OF DEVELOPMENT 

|
|
An e-publication by the World Agroforestry Centre |
|
AGROFORESTRY A DECADE OF DEVELOPMENT |
|
|
section 5 Chapter 15 Y. R. Dommergues Introduction Agroforestry represents an approach to integrated land use involving the more or less intimate association of different plant species, always including trees or woody perennials on the same unit of land (Huxley, 1983). This definition encompasses the association of trees with perennial crops, such as coffee or cacao, pasture species, annual or biennial crops, trees being planted as windbreaks or in alleys and frequently trimmed during cropping, various forms of multispecies combinations such as those found in homegardens, and also mixed forests (mixed perennial cropping). The most decisive factor for the success of agroforestry is the choice of suitable, useable tree species (Nair et al, 1984). This choice should be based on economic and agronomic criteria. In order to fulfil the second criterion, the highest priority should be given to selecting trees that can improve the soil, and identifying species or clones that will fix or absorb large amounts of N (and also other elements, especially P) and then return them to the soil (Huxley, 1983). Nitrogen-fixing tree species are probably the best choice, if they can actively fix nitrogen and thus significantly contribute to the improvement of the nitrogen status of the soil. Nitrogen-fixing trees can increase the yield of associated crops (annual or perennial) through different mechanisms:
In addition, a series of indirect mechanisms (i) affect soil physical, chemical and biological properties, such as structure (Sprent, 1983), or nitrification, whose reduced intensity alleviates the inhibiting effect of nitrates on nitrogen fixation; (ii) control diseases or pests, such as root nematodes (Mulongoy, personal communication); or (iii) stimulate the activity of beneficial organisms such as earthworms (Wilson et al, 1986). In this chapter the discussion is restricted to the nitrogen-fixation process, starting with the most promising nitrogen-fixing species for the tropics, and continuing with current management techniques, and trends and directions for future agroforestry research.
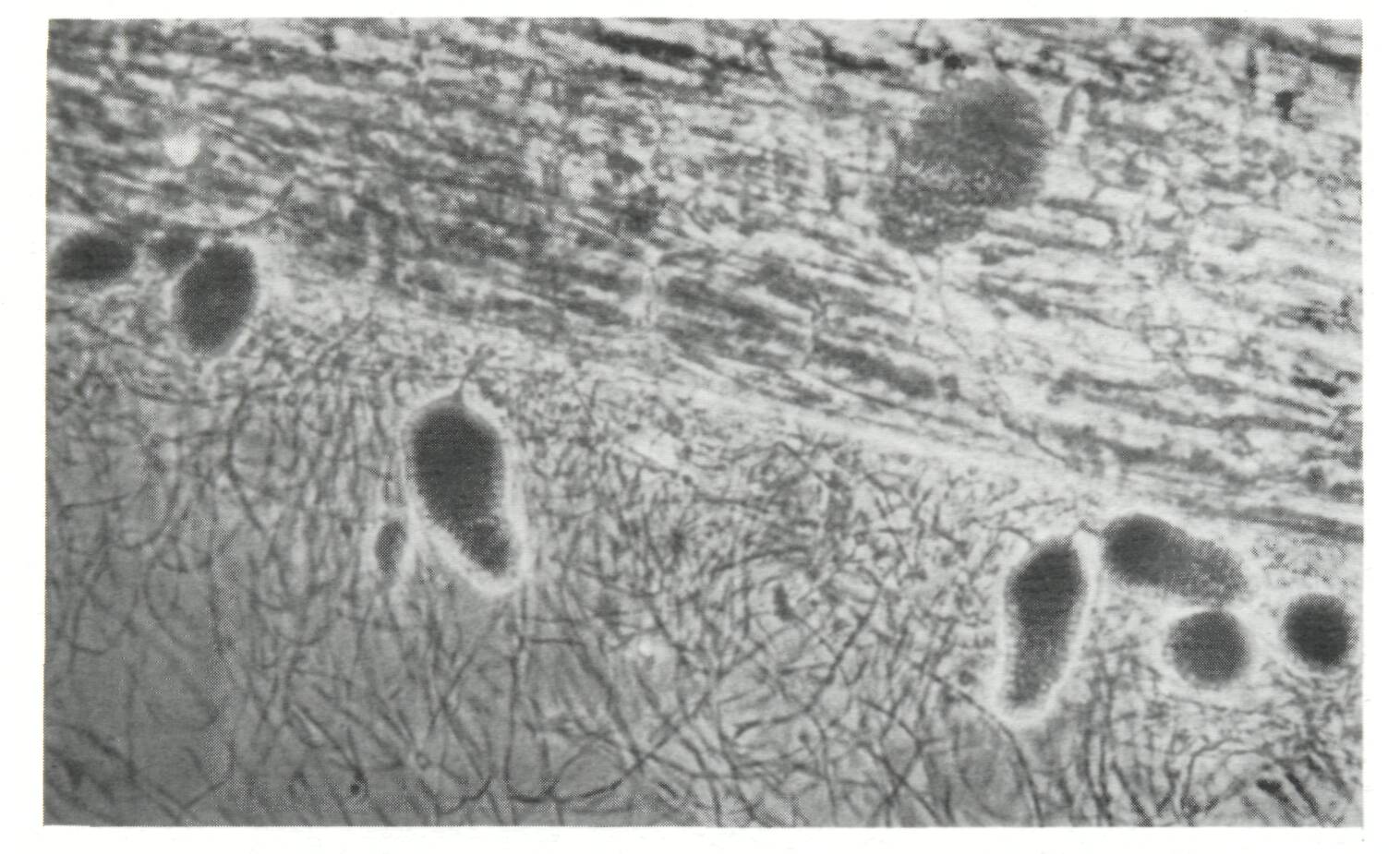
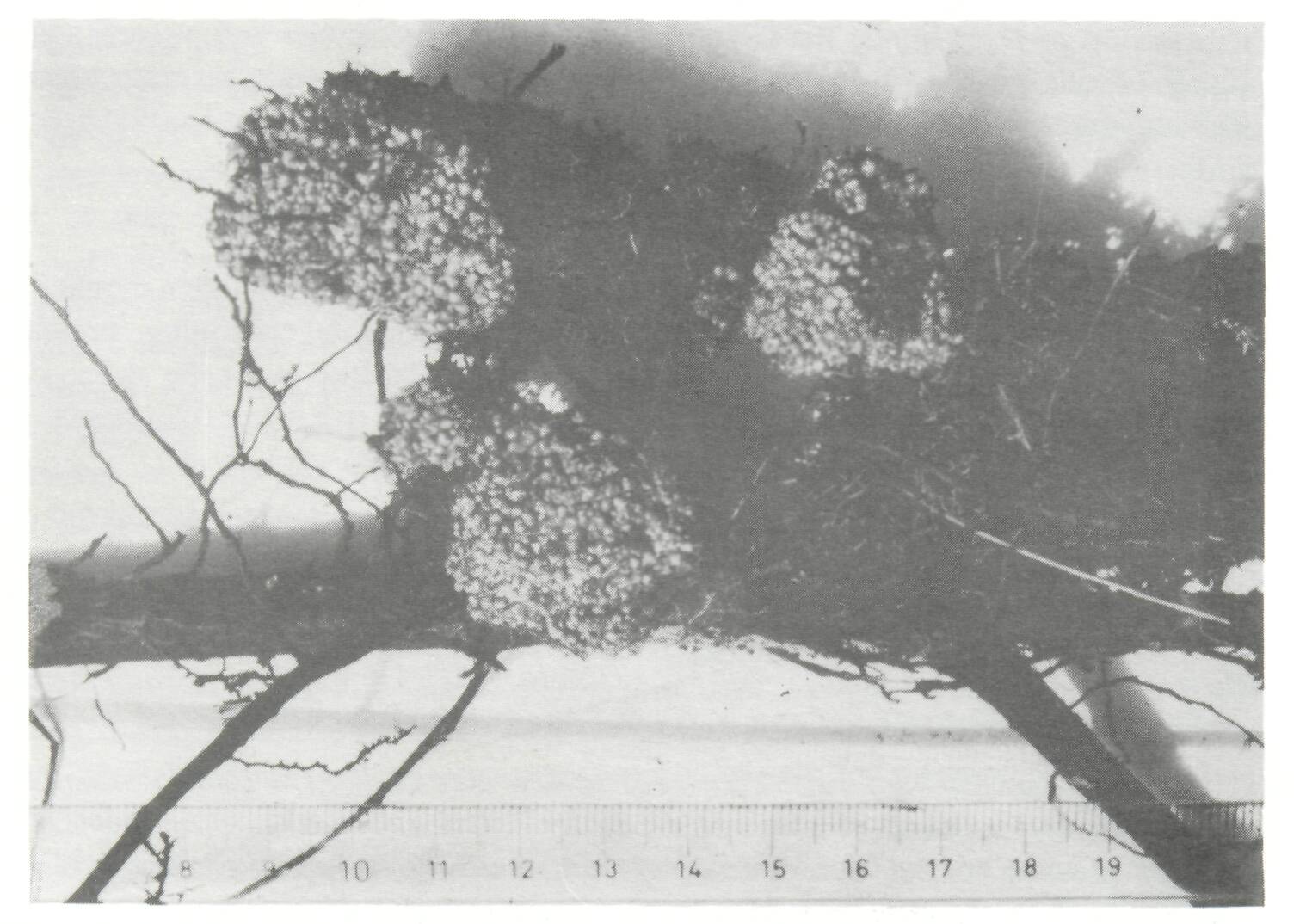
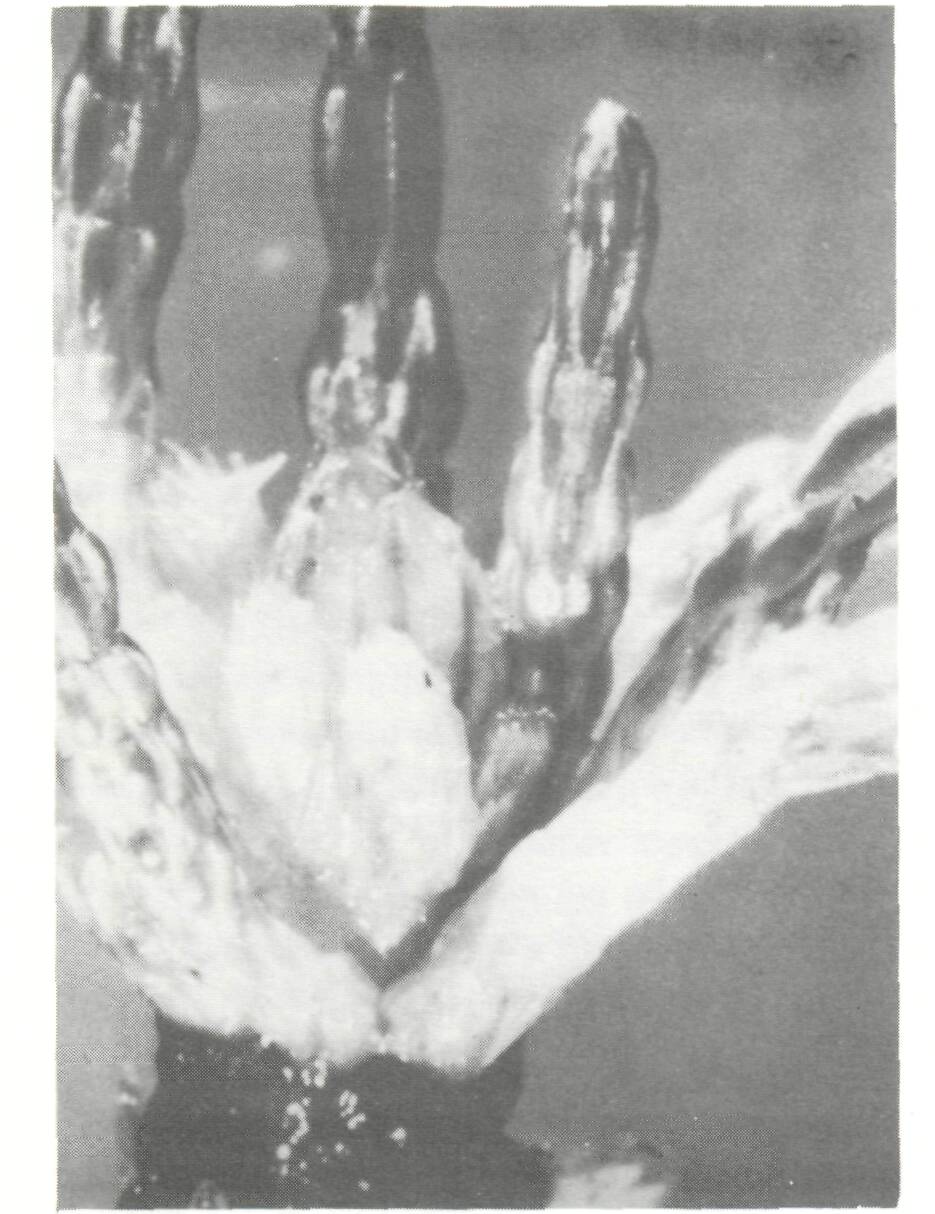
Legumes Only nodulating, that is potentially nitrogen-fixing, legume trees are dealt with in this section. No attention is given to species such as Parkia biglobosa or Cassia siamea which do not nodulate, and consequently do not fix nitrogen, but are nevertheless, sometimes integrated in agroforestry systems. Tropical legumes may nodulate with two types of Rhizobium: fast-growing strains which belong to the genus Rhizobium (sensu stricto), and slow-growing strains which form the cowpea miscellany and are now designated as Bradyrhizobium (Elkan, 1984). One category of trees nodulates only with Rhizobium (sensu stricto), e.g., Leucaena leucocephala (with a few exceptions) or Sesbania grandiflora. Another category nodulates only with Bradyrhizobium, e.g., Acacia meamsii or A. albida. A third category is more promiscuous since it nodulates both with Rhizobium and Bradyrhizobium, e.g., Acacia seyal. Acacia Acacia albida (syn. Faidherbia albida) This tree, native to Africa, is usually considered as a highly valuable component of agroforestry systems, not only as a soil improver, but also as a source of fuelwood and forage in semi-arid zones, especially in Africa (Giffard, 1964, 1971; Felker, 1978; Charreau, 1985; Le Houerou, 1985; Poschen, 1986; CTFT, 1986). The yield of crops such as sorghum, peanut, or maize grown under a tree canopy of A. albida is, on the average, substantially higher than when grown alone, which indicates that the tree improves the soil fertility. However, it is not known whether the beneficial effect is due to nitrogen fixation or to other mechanisms enumerated in the introduction. Acacia albida nodulates with strains of Bradyrhizobium (Dreyfus and Dommergues, 1981). Since these strains are already present in most soils, A. albida can be expected to respond poorly to inoculation except in sterilized nursery soils. Nodulation is normally observed on young seedlings, but nodules are seldom found in the field, which suggests that the nitrogen-fixing potential of this Acacia is rather low. However, this conclusion requires confirmation through precise measurements. The nitrogen-fixing potential of A. albida could possibly be improved by capitalizing on its great genetic variability. Acacia Senegal This is another African Acacia species which is intercropped with food crops in the semi-arid zone (El Houri Ahmed, 1979). A. Senegal nodulates only with fast-growing strains of Rhizobium, i.e., Rhizobium (sensu stricto) (Dreyfus and Dommergues, 1981). Since these strains are less ubiquitous than strains of Bradyrhizobium, one can predict that Acacia Senegal will require inoculation more often than A. albida. The nitrogen-fixing potential of A. Senegal has not yet been estimated. Acacia nilotica, A. raddiana (syn. A. tortilis) and A. seyal Like A. albida and A. Senegal, these acacias are often integrated into different types of agroforestry system in the semi-arid zones. A. nilotica and A. raddiana nodulate with fast-growing Rhizobium whereas A. seyal nodulates with both fast- (Rhizobium, sensu stricto) and slow-growing (Bradyrhizobium) strains (Dreyfus and Dommergues, 1981). The nitrogen-fixing potential of the three species is still unknown. Acacia auriculiformis and A. mangium A. auriculiformis has been reported to be used in agroforestry fuelwood production systems in Papua New Guinea (Nair et al, 1984). It produces profuse bundles of nodules, which suggests a good nitrogen-fixing potential (Domingo, 1983). Acacia mangium hybridizes naturally with its close relative A. auriculiformis (National Research Council, 1983). A. mangium is assumed to be a good nitrogen fixer and, hence, a promising component in fuelwood production systems when grown in appropriate climatic zones. Acacia meamsii (syn. A. mollissima or A. decurrens var. mollissima) A highland tree native to Australia, A. meamsii has been introduced in many countries for its bark which is very rich in tannins. It is also frequently found as a multipurpose tree on farmlands in Java and eastern Africa (Nair et al., 1984). A. meamsii nodulates profusely with strains of Bradyrhizobium (Halliday and Somasegaran, 1982), even in very poor soils, provided the pH is not lower than 4.5. Nitrogen fixation is high: it was estimated to be approximately 200 kg N2 ha-1 yr-1 by Orchard and Darby (1956). A similar figure was given recently by Wiersum (1985). Albizia lebbeck and A. falcataria There are about 100 species of Albizia distributed throughout Africa, Asia and tropical America, Two species, Albizia lebbeck and A. falcataria, are ideally adapted to agroforestry systems; they are renowned as soil improvers because of their profuse nodulation. Native to India, Bangladesh, Burma and Pakistan, Albizia lebbeck is widely cultivated in tropical and subtropical regions with an annual rainfall as divergent as 500 and 2,000 mm (National Research Council, 1979). Perennial nodules of large size have been observed on adult trees in Senegal. Native to the eastern islands of the Indonesian archipelago and the west of Irian, Albizia falcataria (syn. Paraserianthes falcataria) has been spread throughout South-East Asia. It is one of the fastest growing trees in the world provided that it is grown on sites where annual rainfall is high enough (2,000-2,500 mm) (National Research Council, 1979). It nodulates abundantly, which suggests a good nitrogen-fixing capacity. However, because it is exacting in its soil requirements, A. falcataria is probably a poor nitrogen-fixer when it is introduced in relatively infertile soils. Calliandra calothyrsus Native to Central America, this legume was introduced in Indonesia in 1936 with seeds from Guatemala, and has been shown to nodulate with a fast-growing strain of Rhizobium (Halliday and Somasegaran, 1982). Although prized as a first-class soil improver in rotation schedules and in intercropped systems (Domingo, 1983; Nair et at, 1984), considerably more research is needed to accurately assess its nitrogen-fixing potential and its use as forage and organic fertilizer (Baggio and Heuveldop, 1984). Erythrina spp. More than 100 species are planted as shade trees, windbreaks, living fences, plant supports, for food and even medicinal purposes, or as components of alley-cropping systems (NFTA, 1986). E poeppigiana, a fast-growing species, is widely used for shade in coffee and cacao plantations (Budowski, this volume). It has been shown to nodulate with a strain of Bradyrhizobium (Halliday and Somasegaran, 1982). Nodules of E poeppigiana tend to be large, spherical, and clustered on the central root system (Allen and Allen, 1981). The biomass of the root nodules varied from 80 to 205 mg (dry weight) dm-3 soil, being highest close to the stem of the tree (Lindblad and Russo, 1986). A conservative estimate made in Venezuela and based on the decomposition of nodules during the dry season, indicated that the rate of nitrogen fixation was approximately 60 kg N2 ha-1 yr-1 (Escalante et al, 1984). Gliricidia sepium Native to Central America, this species is widely used for shading cacao (Mexico) or coffee (Sri Lanka) plantations. It has been shown to nodulate with fast-growing Rhizobium (Halliday and Somasegaran, 1982). Further experiments are needed to confirm this observation. Estimates of nitrogen fixation based on nodule biomass and rates of nitrogenase activity are approximately 13 kg N2 ha-1 yr-1 in the conditions prevailing in Mexico (Roskoski et al, 1982). G. sepium has been introduced in western Africa in alley-cropping systems (Wilson et al, 1986; Kang and Wilson, this volume) (Figure 1), but its nitrogen-fixing activity may be impeded by attacks of root nematodes. Sumberg (1985) reports that different accessions of G. sepium exhibit considerable variation. This large genetic variability should be exploited to improve nitrogen fixation. Inga jinicuil This species, often found in the same sites as Inga vera, is a popular shade tree in coffee plantations in Mexico. In a plantation in Xalapa, annual nitrogen-fixation rates, based on the acetylene reduction method, were 35-40 kg N2 ha-1 yr-1, which, when compared to nitrogen from fertilizers, represents an important nitrogen input. The corresponding nodule biomass was 71 ± 14 kg (dry matter) ha-1. Given a density of 205 trees ha-1, nodule biomass per tree was 346 g (dry weight), a figure similar to that reported by Akkermans and Houvers (1983) for Alnus (Roskoski, 1981 and 1982) (see Figure 2). Mimosa scabrella Native to the Parana region of south-east Brazil, Mimosa scabrella is used in humid and subhumid tropical highlands as a multipurpose tree on farmlands and as a fuelwood producer in agroforestry operations (Nair et al, 1984). It was shown to nodulate with a fast-growing strain of Rhizobium (Halliday and Samosegaran, 1982). Mimosa scabrella responds positively to inoculation (Dobereiner, 1984), but its exact nitrogen-fixing potential has not yet been evaluated. Leucaena leucocephala This tree has been the focus of a great deal of research in the past few decades (National Research Council, 1977; IDRC, 1983; Brewbaker, this volume). Native to Central America, it has been planted in many tropical countries, including south-east Asia (Domingo, 1983), Africa (Okigbo, 1984; Sanginga et al., 1986) and South America (Dobereiner, 1984), as a shade tree for commercial crops, alley cropping or agroforestry wood production (Figure 3). The acetylene reduction method (Hogberg and Kvarnstrom, 1982) and the difference method (Sanginga et al., 1985, 1986), which have been used to evaluate nitrogen fixation by Leucaena leucocephala, give figures in the range of 100-500 kg N2 ha-1 yr-1. These figures have been confirmed recently by Sanginga, Mulungoy and Ayanaba (personal communication), who used the 15N dilution method to make a precision evaluation of the nitrogen-fixation rate of Leucaena leucocephala grown in an Alfisol, pH 6.1, at the International Institute of Tropical Agriculture (IITA) in Ibadan, Nigeria. They showed that Leucaena leucocephala fixed 98-134 kg N2 ha-1 in 6 months. The high nitrogen-fixing potential of this tree is related to its abundant nodulation under specific soil conditions, in which the nodule dry weight was reported to reach approximately 51 kg ha-1 in a stand of 830 trees ha-1 (Hogberg and Kvarnstrom, 1982), and approximately 63 kg ha-1 in a stand of 2,500 trees ha-1 (Lulandala and Hall, 1986). Leucaena leucocephala generally nodulates with Rhizobium (sensu stricto) (Halliday and Somasegaran, 1982), and occasionally nodulates with Bradyrhizobium (Dreyfus and Dommergues, 1981). The Rhizobium strain specific to Leucaena leucocephala is not generally found in soils. This explains the positive response to inoculation obtained in most soils where the level of nutrients (other than nitrogen) is high enough to satisfy the-tree's requirements. Leucaena leucocephala is not a miracle tree. Its sensitivity to soil acidity and its high nutrient demand are reflected in its poor performance in infertile soils, e.g., the sandy soils of the Pointe Noire region of the Congo or of Hainan Island, China, even when properly inoculated and grown in a suitable climate. Sesbania grandiflora Native to Asia, this legume tree is popular throughout South and South-East Asia, where it is used as a shade tree, a source of fodder and green manure, and for erosion control. Like other Sesbania, Sesbania grandiflora nodules with fast-growing strains of Rhizobium (Dreyfus, personal communication). It nodulates profusely and is probably a good nitrogen fixer (Domingo, 1983), but we have observed that in some soils (e.g., Loudima, Congo) its root system reacted badly to nematode attacks. Actinorhizal plants About 200 non-leguminous plant species belonging to 19 genera and 8 families nodulate with N2-fixing micro-organisms known as Frankia (Figure 4). Since Frankia are actinomycetes, these N2-fixing plants became known as "actinorhizal plants" (Torrey and Tjepkema, 1979), a name now used world wide. In tropical agroforestry the main species of actinorhizal plants belong to the genera Alnus, Casuarina and Allocasuarina and, secondarily, Coriaria (Akkermans and Houvers, 1983; Gauthier et al, 1984; Bond, 1983). Alnus jorullensis (syn. A. acuminata) In Costa Rica, an agroforestry system composed of this actinorhizal species and pasture grasses has become very popular, probably because of the high nitrogen-fixing potential of Alnus (Budowski, 1983). The actual amount of nitrogen fixed and transferred to the pasture is unknown. Introduction of A. jorullensis into coffee plantations has been advocated by Fournier (1979). The great genetic variability of Alnus jorullensis can be exploited to enhance the nitrogen-fixing potential of this species. Casuarinaceae The family Casuarinaceae consists of a group of 82 species mostly from Australia, but also native to South-East Asia and the Pacific islands. Johnson (1982) recognizes four genera: Casuarina (sensu stricto) (e.g., C. cunninghamiana, C. equisetifolia, C. junghuhniana syn. C. montana, C. glauca, C. obesa, C. oligodori); Allocasuarina (e.g., A. decaisneana, A. fraseriana, A. littoralis, A. torulosa, A. stricta syn. Casuarina verticillata); Gymnostoma (e.g., G. deplancheana, G. papuana, G. mmphiana), and a fourth genus not yet described. In some parts of the world, Casuarina spp. are perfectly integrated in agroforestry systems. One classical example is that of Papua New Guinea, where C. oligodon and G. papuana (in the highlands) and C. equisetifolia (in the lowlands) are intercropped with food crops, and are used as shade trees or in rotation with crops (Thiagalingam, 1983; Bourke, 1985). In India, Casuarina equisetifolia plantations are associated with crops such as peanuts, sesame and various grain legumes (pulses) (Kondas, 1981). In many places casuarinas are planted as windbreaks: C. glauca in Tunisia (Figure 5) and E. equisetifolia in Senegal (protection of market-gardens) and Corsica (protection of cash crops such as Actinidia sinensis). According to Bowen and Reddel (1986), Casuarina (sensu stricto) are usually well nodulated, whereas nodulation of Allocasuarina is variable or often non-existent. There is little cross-inoculation between Casuarina and Allocasuarina, which means that strains of Frankia isolated from Casuarina do not usually infect Allocasuarina, and vice versa. The dry weight of a Casuarina equisetifolia nodule in the field is comparable or even greater than that of Alnus (up to 500 g for a 13-year old Casuarina equisetifolia, according to Cao Yue Hua, personal communication) (Figure 6). This profuse nodulation explains its high nitrogen-fixing potential. In an experiment conducted in irrigated microplots at the ORSTOM Station in Dakar, Senegal, a selected clone of Casuarina equisetifolia was reported to fix 42.5 g per tree during the first nine months following plantation. Extrapolating this value to 1 ha with 2,000 trees, and assuming a constant fixation rate, would give a figure of 113 kg N2 fixed per hectare during the first year (Sougoufara, personal communication). The nitrogen-fixation estimate for a 13-year rainfed Casuarina equisetifolia plantation located on sand dunes along the Senegalese coast was only 58 kg N2 ha-1 yr-1 (Dommergues, 1963).
Some Casuarina species are good candidates for the reclamation of salt-affected soils. Recently, Bowen's group in Australia showed that C. obesa inoculated with a salt-tolerant strain of Frankia grew despite 15,000 ppm chloride (approximately the equivalent of sea water), thus tolerating ten times more chloride than when it grew on nitrate or ammonium nitrogen. This is possibly because the nitrogen-fixing system is considerably less salt-sensitive than the mechanism for uptake of nitrate or ammonium, and/or assimilation enzymes (Bowen and Rosbrook, 1986). Coriaria All the 15 species of Coriaria are recorded as bearing nodules, which indicates that nodulation is a generic character of Coriaria, as it is in Alnus. Two species are known to be valuable components in agroforestry systems. One is C. sinica, a deciduous bush, widely grown in Hunan, China, as a source of green manure and of feed for silkworms. It grows so fast that the stems can be harvested 4-5 times a year giving 10t of fresh biomass ha-1 (Watanabe, personal communication). The other is Coriaria arborea, which, grown as an understorey species in many artificial plantations of Pinns radiata in New Zealand, fixes up to 192 kg N2 ha-1 yr-1. Its effect on the growth of Pinus radiata has not yet been investigated (Silvester, 1977, 1983).
This section presents briefly the principles for choosing the species and provenances of nitrogen-fixing trees and describes the practices that are currently recommended to alleviate some of the major environmental stresses inhibiting the nitrogen-fixation process. Choosing species and provenances of nitrogen-fixing trees The species or provenances chosen for introduction in any agroforestry system should have the highest nitrogen-fixing potential, be tolerant of environmental constraints, especially low levels of nutrients, and resistant to pests. Results obtained from the sparse data available on the nitrogen-fixing potential of trees suggest that tree species can be put into two broad categories:
Species in the first category can be divided into two subgroups:
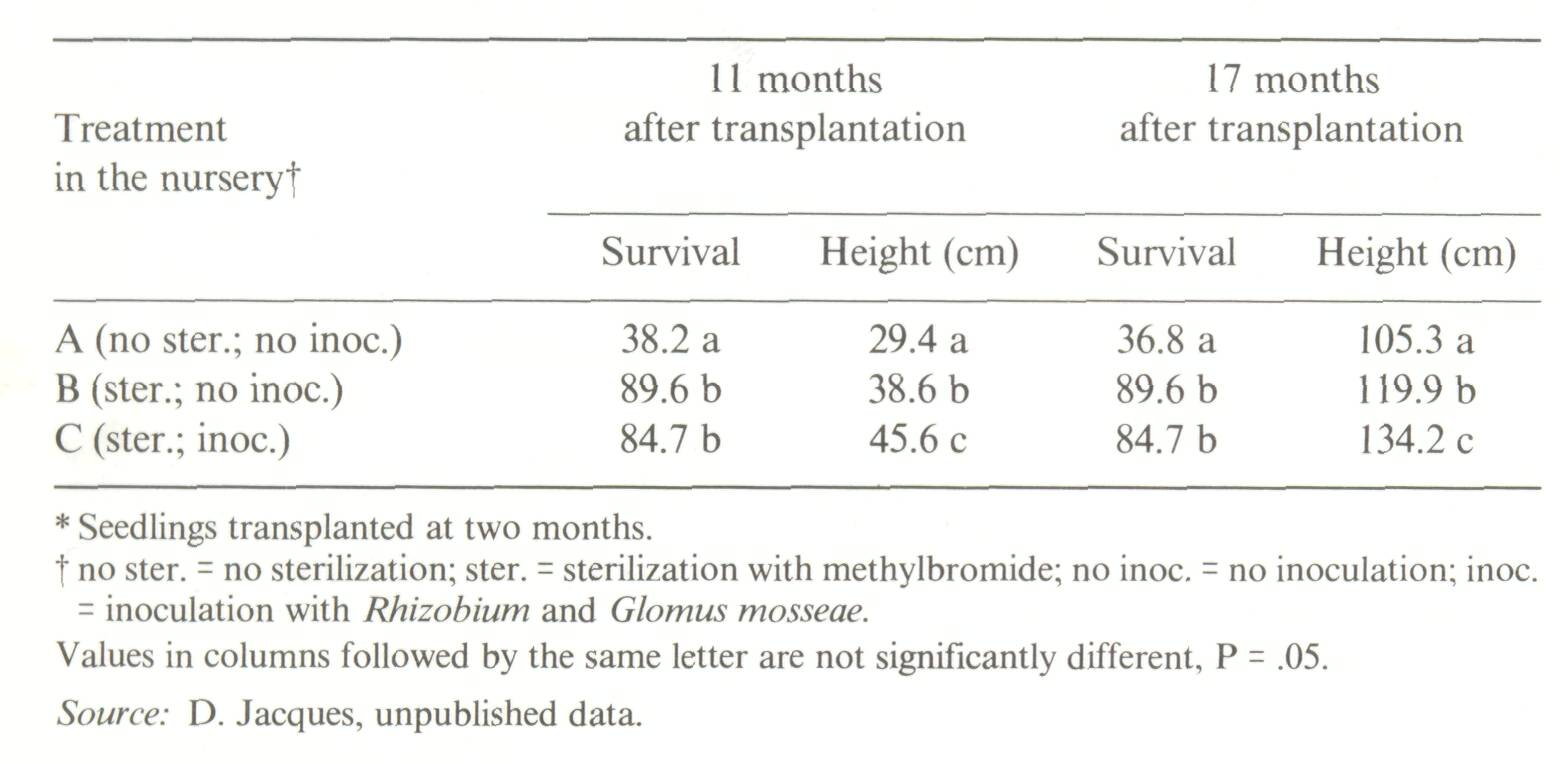
Species belonging to the latter subgroup are obviously the most promising in agroforestry, if they also exhibit the specific qualities required for inclusion in agroforestry systems. The choice of suitable nitrogen-fixing trees for a given agroforestry programme should always be based on well-planned provenance1 trials. The objective of these trials should be to screen as many provenances within species as possible in order to identify provenances best suited to the range of soils and climate existing in a region. Although provenance trials for nitrogen-fixing trees are just beginning, studies are being made to explore provenance variations of species such as Casuarina equisetifolia in China, the Philippines (Halos, 1983), Senegal (Sougoufara and Corbasson, personal communication), or Acacia in Indonesia, Papua New Guinea and Australia (Turnbull et al, 1984), and Congo (D. Diangana and J.C. Delwaulle, personal communications). The choice of the tree species should also be made taking into account its susceptibility to pathogens and pests such as termites (Mitchell et al, 1986) and, especially, nematodes. Introducing tree species whose root systems serve as reservoirs for pathogenic nematodes (Taylor, 1976) should be avoided because of the risk of contamination of associated crops. Sterilization of nursery soils There is a trend towards the generalized use of containers, especially polybags, in many forest nurseries for most indigenous as well as exotic species (Khan, 1985). Taking into account the ease of handling these containers, the limited amount of soil (or any other type of substratum) that is needed for each container, and the relatively low number (2,000-10,000) of plants required per hectare in forest plantations (compared to up to 100,000 ha-1 for annual crops), it is economically feasible to sterilize the soil used to fill the containers. A corollary to sterilization is the necessity to inoculate the containerized seedlings with the appropriate strains of Rhizobium, Frankia or vesicular-arbuscular mycorrhizal fungi. In many tropical soils, pathogens can only be eliminated if the soil in the containers is sterilized. The following example illustrates the spectacular effect of sterilization. Acacia holosericea seedlings were grown in polybags filled up with non-sterilized soil (treatment A), or with sterilized soil. One part of the plants grown in sterilized soil remained uninoculated (treatment B); the other part grown in sterilized soil was inoculated with Rhizobium and a vesicular-arbuscular mycorrhizal fungus, Glomus mosseae (treatment C). When transplanted to the field, trees which had been grown in non-sterilized soil (treatment A) exhibited a survival rate which was less than half that of the plants grown in sterilized soils (treatments B and Q. The effect of inoculation with Rhizobium and Glomus mosseae on height was much less spectacular, but still significant (Table 1). Sterilization of nursery soils is a common practice in temperate countries, but is rarely done in the tropics, probably because the prescribed methods are too expensive (vapour), or dangerous (methylbromide), or because the chemical product is difficult to obtain. Dozomet,3 a granulated biocide, which decomposes into gaseous compounds (methylisothiocyanate and formaldehyde) in the soil, leaving no lingering toxic residual effect, can be recommended because it is easy to handle and not too expensive. Its efficiency as a sterilizing agent is satisfactory provided that its rate and duration of application are calculated for the prevailing climatic and edaphic conditions.
Inoculating the host plant with soil or crushed nodules is a technique that is still recommended but should be discontinued because of the high risk of contaminating seedlings or cuttings with root pathogenic agents, such as Rhizoctonia solani or Pseudomonas solanacearum in the case of Casuarina equisetifolia (Liang Zichao, 1986), or nematodes in the case of Australian Acacia introduced in western Africa. In the past, pure cultures of Frankia have not often been used on actinorhizal plants because of the difficulty of isolating and cultivating the strains, especially those of Casuarina (Diem et al., 1982, 1983), and consequently of obtaining the inoculants. However, thanks to recent progress in knowledge of Frankia physiology, there is reason to hope that actinorhizal plants will be inoculated with pure cultures of Frankia in the near future. Von Carlowitz (1986) has drawn up a list of suppliers of strains or inoculants, most of which still have to be evaluated for nodulation or nitrogen fixation (Somasegaran and Hoben, 1985). In the case of trees raised in containers, inoculation with Rhizobium is best achieved by spraying or drilling the inoculum directly into the container at the time of seeding or planting. When dealing with Frankia, it is advisable to mix the soil or substratum of the container with the inoculum because Frankia, like vesicular-arbuscular mycorrhizal fungi, is not mobile in the soil (Figure 7). After the containerized plants have been transplanted to the field, the effect of inoculation observed in the sterile nursery soil persists only if the soil does not contain specific native strains. Pre-inoculated seeds of Leucaena leucocephala were sown directly in the field by Sanginga et al. (1986) who tested IRc 1045 and IRc 1050 Rhizobium strains in agroforestry experiments set up at two locations in Nigeria. At both places, inoculated trees produced more nitrogen and dry matter than the controls. This effect was statistically equivalent to the application of 150 kg ha-1 of urea. Further, the strains survived and competed well in the field, as was shown in observations made one year after their establishment. The beneficial effect of Leucaena leucocephala on a subsequent crop of maize is discussed later in this chapter. Inoculation with mycorrhizal fungi Mycorrhizal infections are known to increase the absorption of phosphate and other poorly mobile ions in soil such as Zn, Cu, Mo, and K. Mycorrhizal fungi are most often associated with the roots of nitrogen-fixing trees, endomycorrhizae being more frequent than ectomycorrhizae, the other major type of mycorrhizae. Nodulation and nitrogen fixation require a high P status in the host plant, which can be facilitated by the mycorrhizal symbiont. The beneficial effect of mycorrhizal infection on nitrogen fixation is similar to that of added P in P-deficient soils. Mycorrhizae can enhance the effects of even a small amount of P fertilizer that is added to soils with a serious P-deficiency (Ganry et al, 1985). In addition to improving nutrient absorption, mycorrhizal fungi also affect the physiology of the host plant, enabling it to increase its water uptake, improve its hormone balance and overcome the first-year dormancy of cuttings (Hayman, 1986). The technology of inoculation with ectomycorrhizae is now fully applicable (Schenck, 1982). For endomycorrhizal fungi promising results have already been reported in forest nurseries (Cornet et al., 1982), but the technology is not yet ready for extension to the small farmer. Fertilizer There is a tendency to neglect the mineral nutrition of nitrogen-fixing trees. This is most irresponsible when dealing with exacting species such as Leucaena leucocephala whose exceptional capacity to produce biomass and protein depends on the availability of adequate nutrients (Waring, 1985). Hu and Kiang (1983) estimated the nutrient uptake of a three-year-old plantation of Leucaena leucocephala as being P 11-27, K 174-331, Ca 138-305 and Mg 31-62 kg ha-1. These figures are indeed high. Casuarina equisetifolia is assumed to have high Ca requirements (Waring, 1985). P is also an important nutrient, not only for the plant itself but also to ensure good nodulation. However, whether a low P supply blocks nodulation by limiting plant growth and hence nitrogen demand, or directly affects Frankia in the rhizosphere and in the early stages of nodule initiation is not known (Reddell et al, 1986). The nutrient requirements of species such as Acacia mangium that are less constrained by element deficiency are probably lower but not low enough to be negligible. It is surprising that some authors still recommend the application of nitrogen fertilizers (together with P and K) on nitrogen-fixing trees (Yadav, 1983). This practice should be prohibited, since it is a well-established fact that mineral nitrogen, especially when applied at high levels, inhibits nodulation and nitrogen fixation. Obviously, much more research is necessary to quantify the exact fertilizer needs of nitrogen-fixing trees. As suggested by Sanchez and Salinas (1981), research along these lines should keep "inputs to a minimum, aiming only to optimize production under existing constraints rather than to maximize the production per se". Control of acidity Soil acidity and related factors (Al and/ or Mn toxicity and Ca and Mo deficiencies), which affect many tropical soils (Franco, 1984), influence nitrogen fixation by the direct or indirect effects they have on the host plant and the symbiotic micro-organisms. A typical example is Acacia meamsii, which does not nodulate in the highlands of Burundi where soils have a low pH and a high content of exchangeable Al. The detrimental effects of soil acidity can be overcome by selecting acid-tolerant host plants and symbiotic microorganisms, an approach that has been adopted with Leucaena leucocephala (Hutton, 1984; Brewbaker, this volume) and its competent Rhizobium (Halliday and Somasegaran, 1982; Franco, 1984). It is also possible to control the effects of soil acidity by directly applying proper amendments to the soil or by pelleting the seeds in the case of direct sowing in the field. Different types of amendments such as lime or organic materials can be used. The acidity generated by nitrogen-fixing plants in the long run may lower the pH of weakly buffered soils, and periodic liming may be necessary to maintain high productivity (Franco, 1984). The higher organic-matter content of soil under nitrogen-fixing trees, however, may lead to satisfactory yields even when the pH is lower than usually recommended in conventional cropping systems. The symbiotic micro-organism can be protected against acidity by pelleting the seeds to be inoculated with calcium carbonate or rock phosphate. This technique, developed in Australia and now used throughout the world, has indeed proved to be a high-value alternative for liming during the introduction and establishment of forage legumes in pastures (Williams, 1984). It could also be used successfully in agroforestry. However, in very acid soils with Al or Mn toxicity, pelleting the seeds alone cannot overcome the effects of acidity regardless of cropping system.
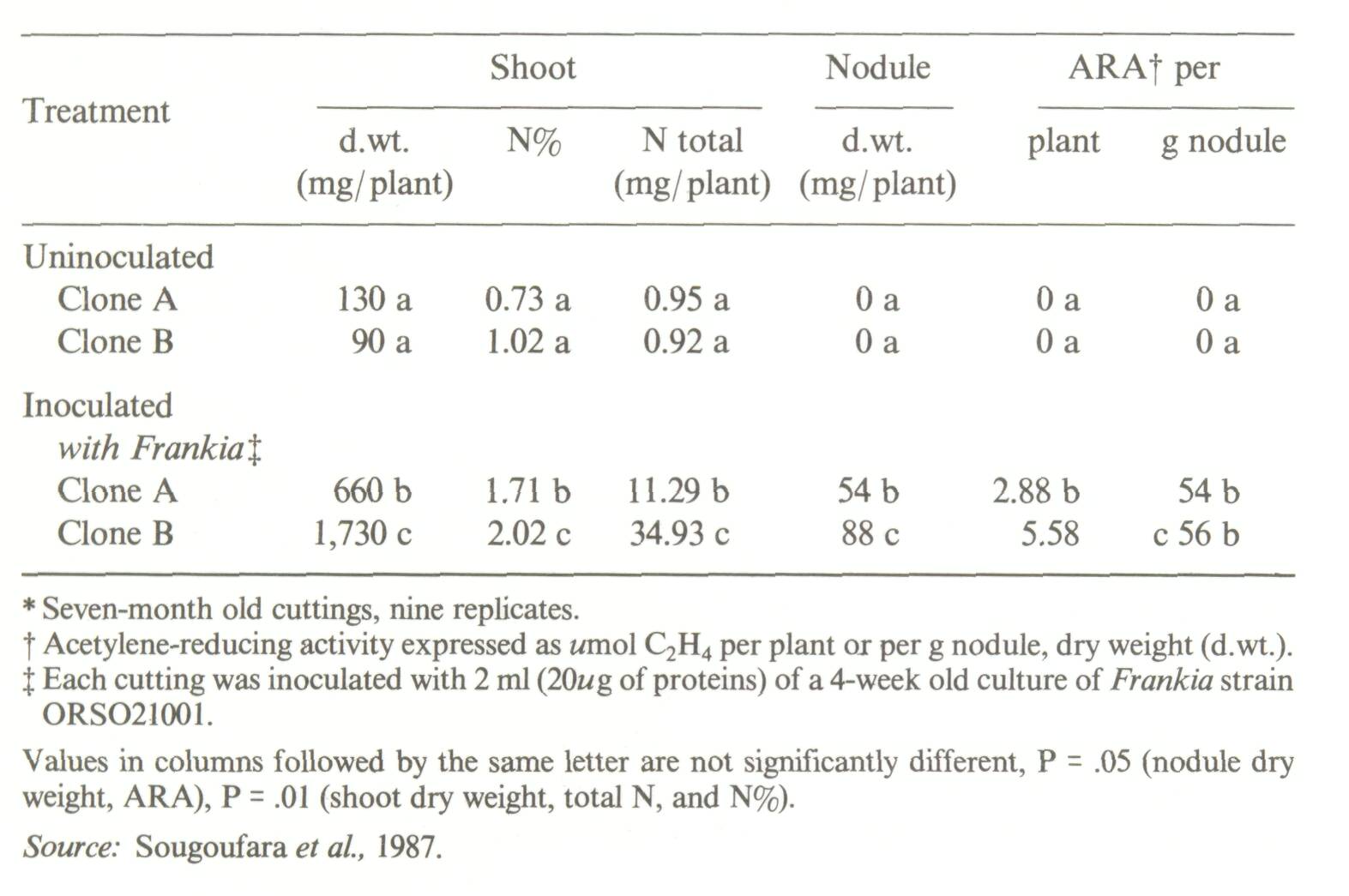
In the last decade a tremendous amount of investigation has been devoted to the genetics and physiology of Rhizobium, but until recently forestry and agroforestry had not benefited substantially from the remarkable progress that has been achieved in this field. Hopefully some of the encouraging results that have already been obtained will be transferred to the field. In addition, new concepts and techniques are presently emerging that will, sooner or later, be ready for practical application. The most promising areas of investigation are probably those concerned with improvement of the host plant. Improvement of the symbiotic micro-organism Near-term investigations To date only relatively few effective strains of Rhizobium that nodulate nitrogen-fixing trees have been isolated: some of the best known are strains for Leucaena leucocephala, e.g., strain TAL1145 from NifTAL2 (Roskoski, 1986) and strains IRc 1045 and 1050 from IITA (Sanginga et al, 1986). There is still much work ahead to collect Rhizobium strains for leguminous nitrogen-fixing trees and then screen them for genetic compatibility, nitrogen-fixation effectiveness, and tolerance to environmental stresses, especially soil acidity (Da Silva and Franco, 1984). In addition to preliminary screenings, field trials must be performed to test the response to inoculation since "accumulating data indicate that site variation in performance of selected strains is common" (Halliday, 1984a). Frankia strains associated with Casuarinaceae exhibit large differences in genetic compatibility (Zhang and Torrey, 1985; Puppo et al., 1985) and effectiveness (Zhang et al., 1984; Bowen and Rosbrook, 1986). Not all Frankia strains nodulate all species of Casuarinaceae. There are very large differences in the effectiveness of nitrogen fixation between Frankia strains associated with a single species of Casuarinaceae. Furthermore, a Frankia strain effective on one species of Casuarinaceae can be very ineffective on another species (Reddell, 1986). Collections of Frankia strains already exist (Lechevallier, 1985-6), and some laboratories have already screened and selected strains for use in nursery and field trials. Mid- and long-term investigations Using molecular techniques (molecular cloning and recombination), new strains of Rhizobium and Frankia will probably be engineered to contain multiple copies of the major genes involved in the symbiosis: genes of nitrogen fixation and nodulation, and genes involved in interstrain competition. Since Rhizobium and Frankia are exposed to environmental constraints in the field, the new strains should also be stress tolerant. Instead of developing strains with the superior traits indicated above and then studying their behaviour in the field, "bacteria whose genetic libraries already contain adaptation traits to prevailing environmental stresses will be engineered for better nodulation and nitrogen fixation" (Roskoski, 1986) and competitive ability. Improvement of the host plant The amount of nitrogen fixed by any nitrogen-fixing tree is related to its nitrogen-fixing potential (NFP), i.e., its ability to-fix nitrogen in the absence of any limiting factor (Halliday, 1984b). The nitrogen-fixing potential is directly conditioned by the genotypes of both the host plant and the associated symbiont. Consequently, to get the maximum nitrogen input into an agroforestry system, the first essential characteristic is to use a nitrogen-fixing tree with a high nitrogen-fixing potential. The second essential characteristic is that a nitrogen-fixing tree should be maximally tolerant of environmental stresses, be they physical (e.g., excessive temperature, drought), chemical (e.g., excess of combined nitrogen), or biological. Because of these stresses, however, even the most tolerant nitrogen-fixing tree cannot attain its full potential in the field. The amount of nitrogen that is fixed under field conditions is called the actual nitrogen fixation (ANF). The ANF of stress-sensitive nitrogen-fixing trees is expected to be much lower than their NFP; conversely the ANF of stress-tolerant species is expected to be much closer to their NFP. Special mention must be made of the inhibitory effect of high levels of combined (mineral) soil nitrogen, especially nitrate, on nitrogen fixation (as mentioned earlier). This implies that nitrogen-fixing trees in agroforestry systems should be engineered to continue fixing significant amounts of nitrogen even when the intercrop receives nitrogen fertilizers. A third group of essential characteristics, common to all trees to be introduced in agroforestry systems, has been presented by Huxley (1983). One trait worth mention is the absence of strong plant competitive attributes such as a tendency to overshade understorey plants and to dominate the water economy of the microsite. Short-term investigations When the best provenances have been identified, it is mandatory to study differences that occur between the individual trees, especially differences hi ANF. Exploiting such spontaneous variations requires two steps. First, the whole population in the provenance must be screened using an adapted procedure to identify the more actively nitrogen-fixing individuals. This procedure might be based on study of the nodulation combined with the measurement of the acetylene-reducing activity of the different individuals. Secondly, the superior phenotypes must be vegetatively propagated, using available techniques (Datta and Datta, 1984; Leaky, 1986; Duhoux et al, 1986). Sougoufarae et al. (1987) recently used this approach successfully. The result was a spectacular increase of the nitrogen-fixing potential of Casuarina equisetifolia (Figure 8). These authors identified a clone of Casuarina equisetifolia (called clone B) with a much higher nitrogen-fixing potential than that of a reference clone (clone A), i.e., a clone with a potential similar to that of the seedlings usually grown. Clones A and B were grown in a sterile nitrogen-deficient soil. One set of clones A and B was inoculated with the same Frankia strain, the other one remained uninoculated. After seven months, the uninoculated clones displayed poor growth, while the inoculated clones had grown satisfactorily, but their response to inoculation differed markedly. Inoculated clone B produced 2.6 times more biomass (expressed in terms of dry weight and total nitrogen) than inoculated clone A. Concomitantly, nodule weight and nitrogen fixation (expressed as acetylene-reducing activity per plant) of clone B were significantly higher (1.6 times) than those of clone A. The difference in the nitrogen-fixing potential of the clones appeared to be related to their nodule weight (Table 2). Field trials under way in Senegal will, hopefully, confirm the superior nitrogen-fixing potential of clone B. Mid- or long-term investigations for improving nitrogen-fixing trees The methods mentioned below can be used either for improving the nitrogen-fixing potential or the stress tolerance, or both traits, of nitrogen-fixing trees.
Field studies on nitrogen fixation Estimation of nitrogen fixation With a few exceptions (Leucaena leucocephala and Casuarina equisetifolia), the NFP or ANF of most nitrogen-fixing trees is not precisely known. This dearth of information is prejudicial to the development of proper management practices. The principles of the current methods have been discussed in many reviews (e.g., LaRue and Patterson, 1981; Herridge, 1982; Silvester, 1983) and described in Bergersen's treatise published in 1980.
In ureide exporters, much of the nitrate absorbed by the roots is passed to the shoot as free unreduced nitrate because of the low nitrate reductase activity of their roots. In non-nitrogen-fixing plants, the xylem-nitrogen is found mainly in the form of nitrate and amino acids, whereas in nitrogen-fixing plants it contains mainly ureide nitrogen and the relative abundance of ureides in sap can be used as an indication of nitrogen activity. By constrast, in amide exporters, only a small proportion of the nitrate absorbed by the roots escapes the reductase system of the roots, hence their sap contains mainly amides regardless of whether they are fixing nitrogen or not. This makes it impossible to use sap analysis for estimating nitrogen fixation in amide-exporting legumes (Bergersen, 1986). Preliminary studies on the composition of the sap of 35 nitrogen-fixing leguminous trees have been carried out at NifTAL by Kessel et al. (1987). Only two species, Acacia meamsii and Sesbania grandiflora, showed a high relative abundance of ureides in the xylem sap (81.5 percent in Acacia meamsii and 78.8 percent in Sesbania grandiflora). For the two species, ureides are the major nitrogen compounds in the sap, and the ureide method could probably be used for measuring their nitrogen-fixing potential. Since citrulline is always the major nitrogenous compound in the xylem sap of Casuarina equisetifolia, regardless of whether it is fixing nitrogen or not, the citrulline content cannot be used as an indicator of nitrogen fixation in Casuarina equisetifolia. However, the abundance of citrulline compared to other nitrogenous compounds (e.g., amides or nitrate) could possibly be used as an indicator of nitrogen fixation (Walsh et al., 1984). In sum. there are a number of techniques available to measure nitrogen fixation. Under carefully controlled conditions each will give reasonable estimates (e.g., Herridge, 1982; Gauthier et al., 1985; Bergersen, 1986). Whenever possible at least two methods should be used simultaneously. However, due attention should be given to the difficulties specific to perennial plants, e.g., logistic and sampling problems, variations in the nitrogen-fixing activity with the age of the trees, or interference by difference processes such as losses and redistribution of nitrogen in the different horizons or compartments of the agroforestry system. Transfer of nitrogen fixed by the trees to associated crops In general, the transfer of nitrogen to non-nitrogen-fixing plants intercropped with nitrogen-fixing trees and the rates of nitrogen turnover have not yet been thoroughly assessed. In one of the first studies on this problem, Sanginga et al. (1986) attempted to quantify the beneficial effect of application of Leucaena leucocephala prunings at the surface of plots grown with maize:
Nitrogen from prunings was less well utilized by maize, probably because leaves and twigs of Leucaena leucocephala released 50 percent of their nitrogen within four weeks through decomposition and most of this nitrogen was rapidly mineralized and leached out. Nitrogen transfer studies are urgently needed to improve current management practices and thus capitalize on the benefits that associated crops can get from nitrogen-fixing trees. Mixed perennial systems Mixed forest communities, in which non-nitrogen-fixing trees or diverse perennials are paired with nitrogen-fixing trees in tropical and subtropical countries, include the following couples: (i) Eucalyptus spp. with a number of nitrogen-fixing plants such as Macrozamia sp., which is an understorey nitrogen-fixing cycad (Halliday and Pate, 1976; Grove et al, 1980), or Daviesia mimosoides, which is a common understorey legume in south-eastern Australia (McColl and Edmonds, 1986), or Aicacia sp. (O'Connell et al., 1979; Lawrie, 1981) or Casuarina equisetifolia (personal observation); (ii) Pinus radiata with Coriaria arborea (Gadgil, 1983; Silvester, 1977,1983); (iii) Vitasparviflora, a species valued for its high-quality wood, with Leucaena leucocephala (Domingo, 1983); (iv) coffee with Inga jinicuil (Roskoski, 1981); (v) coffee or cacao with Albizia falcataria (Domingo, 1983; (vi) Acacia nilotica, a relatively poor nitrogen fixer with Leucaena leucocephala, an active nitrogen fixer (Bhatia and Kapoor, 1984). Similar to what has been observed in the case of nitrogen-fixing trees associated with annual crops, nodules seem to constitute an important source of nitrogen for non-nitrogen-fixing species as well. The interrelation between nitrogen-fixing and non-nitrogen-fixing trees is illustrated by the fact that nodules are often found concentrated at the base of the trunk of the non-nitrogen-fixing plants, e.g., coffee-Inga jinicuil (Roskoski, 1981), and Eucalyptus-Casuarina equiietifolia (personal observation) associations. Two types of silvicultural system which have been proposed in temperate countries (Tarrant, 1983) could be tested in the tropics: (i) a system involving nitrogen-fixing trees in mixed plantations or alternate cropping, and (ii) a system involving nitrogen-fixing plants of no economic value, especially shrubs grown as understorey. There are probably many eligible candidate plants, both leguminous and actinorhizal, in the tropics, but they have not yet been tested. The development of research programmes on experimental mixtures comprising Eucalyptus spp. and Acacia mangium, A. melanoxylonand Casuarina sp. was proposed recently by Waring (1985). Some trials involving Eucalyptus sp. and Acacia are already under way in China (Wang Kwon Ming, personal communication) and in Congo (J.C. Delwaulle, personal communication). Such studies are worth pursuing, because observations increasingly indicate that mixed forests provide more stability than pure stands. However, recommending specific tree mixtures before performing field trials can be hazardous, as was evidenced by the recent unsuccessful attempt to interplant Eucalyptus deglupta and Albizia falcataria in the Philippines; it failed because A.falcataria grows taller and much faster than Eucalyptus deglupta, the associated non-nitrogen-fixing tree (Domingo, 1983).
It is widely recognized that nitrogen-fixing trees and shrubs comprise a group of most promising species for agroforestry systems. However, potential direct and ancillary benefits from their introduction vary greatly depending on tree species, climate, soil, and management practices. Six concepts seem important in optimal utilization of biological nitrogen fixation in agroforestry:
Akkermans, A.D.L. and H. Houvers. 1983. Morphology of nitrogen fixers in forest ecosystems. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems: foundations and applications. The Hague: Nijhoff/Junk. Allen, O.N. and E.K.. Allen. 1981. The Leguminosae. A source book of characteristics, uses, and nodulation. Madison: University of Wisconsin Press. Baggio, A. and J. Heuveldop. 1984. Initial performance of Calliandra calothyrsus Meissm. in live fences. Agroforestry Systems 2: 19-29. Bhatia, N. and P. Kapoor. 1984. Neighbor interaction between Leucaena leucocephala and Acacia nilotica in mixed plantations in Punjab. Leucaena Research Reports 5: 18-9. Bergersen, F.J. 1980. Methods for evaluating nitrogen fixation. New York: John Wiley. .1986. Measurements of dinitrogen fixation. In Biotechnology of nitrogen fixation in the tropics (BIOnifT). Proceedings UNESCO Regional Symposium and Workshop, UPM, Malaysia, 25-29 August 1986 (in press). Bond, G. 1983. Taxonomy and distribution of non-legume nitrogen-fixing systems. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems:foundations and applications. The Hague: Nijhoff/Junk. Bourke, R. M. 1985. Food, coffee and casuarina: an agroforestry system from the Papua New Guinea highlands. Agroforestry Systems 1: 273-9. Bowen, G.D. and P. ReddelL 1986. Nitrogen fixation in Casuarinaceae. In Proceedings of the 18th IUFRO World Congress, Ljubljana, Yugoslavia, September 1986 (in press). Bowen, G.D. and P. A. Rosbrook. 1986. The management of nitrogen fixation by Casuarina. CSIRO and ACIAR (mimeo). Budowski, G. 1983. An attempt to quantify some current agroforestry practices in Costa Rica. In P. A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Carroll, B.J., D.L. McNeil and P.M. Gresshoff. 1985. A supernodulating and nitrate tolerant symbiotic (NTS) soybean mutant. Plant Physiol. 78: 34-40. Charreau, C. 1985. Le role des arbres dans les systemes agraires des regions semi-arides tropicales d'Afrique de FOuest. In Le role des arbres au Sahel. Ottawa: IDRC. Cornet, F., H.G. Diem and Y.R. Dommergues. 1982. Effet de 1 Inoculation avec Glomus mossaea sur la croissance d 'Acacia holosericea en pdpiniere et apres transplantation sur le terrain. In Les mycorhizes: biologie et utilisation Paris: INRA. CTFT. 1986. Faidherbia albida. Centre Technique Forestier Tropical, Nogent-sur-Marne. Da Silva, G.G. and A. A. Franco. 1984. Selection of Rhizobium spp. strains in culture medium for acid soils. Pesq. agropec. bras. Brasilia 19: 169-73. Datta, S.K. and K. Datta. 1984. Clonal multiplication of "elite" trees Leucaena leucocephala through tissue culture. Leucaena Research Reports 5: 22-3. Diem, H.G., D. Gauthier and Y.R. Dommergues. 1982. Isolation of Frankia from nodules of Casuarina equisetifolia. Can. J. Microbiol. 28: 526-30. .1983. An effective strain of Frankia from Casuarina sp. Can. J. Bot. 61: 2815-21. Döbereiner, J. 1984. Nodulation and nitrogen fixation in legume trees. Pesq. Agropec. Bras. 19: 83-90. Domingo, I. 1983. Nitrogen fixation in Southeast Asian forestry research and practice. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems: foundations and applications. The Hague: Nijhoff/Junk. Dommergues, Y.R. 1963. Evaluation du taux de fixation de l'azote dans un sol dunaire roboisd en filao (Casuarina equisetifolia). Agrochimka 105: 179-187. Dreyfus, B.L., and Y.R. Dommergues. 1981. Nodulation of Acacia species by fast- and slow-growing tropical strains. Appl. Environ. Microbiol. 41: 97-9. Dreyfus, B.L., D. Alazard and Y.R. Dommergues. 1984. Stem-nodulating rhizobia. In M.J. Klug and C.A. Reddy (eds.), Current perspectives in microbial ecology. Washington, D.C.: American Society for Microbiology. Duhoux, E., B. Sougoufara and Y.R. Dommergues. 1986. Propagation of Casuarina equisetifolia through axillary buds of immature female inflorescences cultured in vitro. Plant Cett Reports 3:161-4. El Houri Ahmed, A. 1979. Effects of land use on soil characteristics in the Sudan. In H.O. Mongi and P.A. Huxley (eds.), Soils research in agroforestry. Nairobi: ICRAF. Elkan, G.H. 1984. Taxonomy and metabolism of Rhizobium and its genetic relationships. In M. Alexander (ed.), Biological nitrogen fixation, ecology, technology, and physiology. New York: Plenum Press. Escalante, G., R. Herrera and J. Aranguren. 1984. Fijacion de nitrogeno en arboles de sombra (Erythrinapoeppigiana) en cacaotales del norte de Venezuela. Pesq. Agropec. bras. Brasilia 19:223-30. Felker, P. 1978. State of the art: Acacia albida as a complementary permanent intercrop with annual crops. Washington D.C.: USAID. Fournier, L.A. 1979. Alder crops (Alnus jorullensis) in coffee plantations. In Proceedings of Workshop on Agroforestry Systems in Latin America. Turrialba, Costa Rica: CATIE. Franco, A.A. 1984. Nitrogen fixation in trees and soil fertility. Pesq. Agropec. Bras. 19: 253-61. Gadgil, R.L. 1983. Biological nitrogen fixation in forestry research and practice in Australia and New Zealand. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems: foundations and applications. The Hague: Nijhoff/Junk. Ganry, F., H.G. Diem, J. Wey and Y.R. Dommergues. 1985. Inoculation with Glomus mosseae improves N2 fixation by field-grown soybeans. Biol. Pert. Soils 1: 15-23. Gauthier, D.L., H.G. Diem, and Y.R. Dommergues. 1984. Tropical and subtropical actinorhizal plants. Pesq. Agropec. Bras. 19: 119-36. Gauthier, D.L., H.G. Diem, Y.R. Dommergues and F. Ganry. 1985. Assessment of N2 fixation by Casuarina equisetifolia inoculated with FrankiaORSQ21001 using 15N methods. Soil Biol. Biochem. 17: 375-9. Giffard, P.L. 1964. Les possibilites de reboisement en Acacia albida au Senegal. Bois et Forets des Tropiques 95: 21-33. .1971. Recherches sur Acacia albida. Bois et Forets des Tropiques 135: 3-20. Grove, T.S., A.M. O'Connell and N. Malajczuk. 1980. Effects of fire on the growth, nutrient content and rate of nitrogen fixation of the cycad Macrozamia riedlei. Austr. J. Bot. 28: 271-81. Halliday, J. 1984a. Principles of Rhizobium strain selection. In M. Alexander (ed.), Biological nitrogen fixation, ecology, technology, and physiology. New York: Plenum Press. .1984b. Integrated approach to nitrogen-fixing tree germplasm development. Pesq. Agropec. Bras. 19:91-117. Halliday, J. and J.S. Pate. 1976. Symbiotic nitrogen fixation by coralloid roots of the cycad Macrozamia riedlei: Physiological characteristics and ecological significance. Austr. J. Plant Physiol. 3: 349-58. Halliday, J. and P. Somasegaran. 1982. Nodulation, nitrogen fixation, and Rhizobium and strain affinities in the genus Leucaena. In Leucaena research in the Asian-Pacific region. Ottawa: IDRC. Halos, S.C. 1983. Casuarinas in Philippines forest development. In S. J. Midgley, J. W. Turnbull and R.D. Johnston (eds.), Casuarina ecology, management and utilization. Melbourne: CSIRO. Hayman, D.S. 1986. Mycorrhizae of nitrogen-fixing legumes. MIRCEN J. 2: 121-45. Herridge, D.F. 1982. A whole-system approach to quantifying biological nitrogen fixation by legumes and associated gains and losses of nitrogen in agricultural systems. In P.H. Graham and S.C. Harris (eds.), Biological nitrogen fixation technology for tropical agriculture. Cali, Colombia: Centre International de Agriculture Tropical. Hogberg, P. and M. Kvarnstrom. 1982. Nitrogen fixation by the woody legume Leucaena leucocephala. Plant Soil 66: 21-8. Hu, T. W. and T. Kiang. 1983. Leucaena research in Taiwan. In Leucaena research in the Asian-Pacific region. Ottawa: IDRC. Hu, T.W., J.C. Huang and C.C. Young. 1985. The role of nitrogen-fixing trees in soil biology. In J. Burley and J.L. Stewart (eds.), Increasing productivity of multipurpose species. Vienna. IUFRO. Hutton, E.M. 1984. Breeding and selecting Leucaena for acid tropical soils. Pesq. agropec. bras. Brasilia 19: 263-74. Huxley, P.A. 1983. The role of trees in agroforestry: some comments. In P. A. Huxley (ed.), Plant Research and Agroforestry. Nairobi: ICRAF. IDRC. 1983. Leucaena research in the Asian-Pacific region. Ottawa: IDRC. Johnson, L.A.S. 1982. Notes on Casuarinaceae. J. Adelaide Bot. Card. 6: 73-87. Kessel, C. van and P. Nakao. 1986. The use of nitrogen-15-depleted ammonium sulf ate for estimating nitrogen fixation by leguminous trees. Agronomy J. 78: 549-51. Kessel, C. van, P. Nakao, J.P. Roskoski and K. Kevin. 1987. Ureide production by N2-fixing leguminous trees. Soil Biol. Biochem. (in press). Khan, S.A. 1985. Nursery practices. In J. Burley and J.L. Stewart (eds.), Increasing productivity of multipurpose species. Vienna: IUFRO. Knowles, R. 1983. Nitrogen fixation in natural plant communities and soils. In F.J. Bergersen (ed.), Methods for evaluating biological nitrogen fixation. New York: John Wiley. Kondas, S. 1981. Casuarina equisetifolia. A multipurpose cash crop in India. In S.J. Midgley, J.W. Turnbull and R.D. Johnston (eds.), Casuarina ecology, management and utilization. Melbourne: CSIRO. Koslowski, T.T. and P.A. Huxley. 1983. The role of controlled environments in agroforestry research. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Kyle, N.E. and T.L. Righetti. 1985. In vitro micrografting of actinorhizal desert shrubs. In H.J. Evans, P.J. Bottomley and W.E. Newton (eds.), Nitrogen fixation research progress. The Hague: Nijhoff/Junk. LaRue, T.A. and G. Patterson. 1981. How much nitrogen do legumes fix? Adv. Agron. 34: 15-38. Lawrie, A.C. 1981. Nitrogen fixation by native Australian legumes. Australian Journal of Botany 29: 143-57. Leaky, R.R.B. 1986. Cloned tropical hardwoods. Quicker genetic gain. Span 29: 35-7. Lechevallier, M.P. 1985-1986. Catalog of Frankia isolates. The Actinomycetes 19: 131-62. Le Houérou, H.N. 1985. Le r61e des arbres et arbustes dans les paturages saheliens. In Le Role des arbres au Sahel. Ottawa: IDRC-158f. Liang Zichao. 1986. Vegetative propagation and selection of Casuarina for resistance to bacterial wilt. Tropical Forestry (Science and Technology) Guangazhou. 2: 1-6 (in Chinese, summary in English). Lindblad, P. and R. Russo. 1986. C2H2-reduction by Erythrinapoeppigiana in a Costa Rican coffee plantation. Agroforestry Systems 4: 33-7. Lulandala, L.L.L. and J.B. Hall. 1986. Leucaena leucocephala's biological nitrogen fixation: a promising substitute for inorganic nitrogen fertilization in agroforestry systems. In Biotechnology of nitrogen fixation in the tropics (BIOnifT), Proceedings of UNESCO Regional Symposium and Workshop, UPM, Malaysia, 25-29 August 1986 (in press). Lundgren, B. 1979. Research strategy for soils in agroforestry. In H.O. Mongi and P.A. Huxley (eds.), Soils research in agroforestry. Nairobi: ICRAF. McColl, J.G. and R.L. Edmonds. 1986. Acetylene reduction by Diviesia mimosaides under Eucalyptus. Plant Soil 96: 215-24. Mitchell, M., D. Gwaze and H. Stewart. 1986. Termite susceptibility of Australian trees in Zimbabwe. ACIAR Forestry Newsletter 2 (Sept./Oct.): 2. Nair, P.K.R., E.C.M. Fernandes and P.N. Wambugu. 1984. Multipurpose leguminous trees and shrubs for agroforestry. Pesq. Agropec. Bras. 19: 295-313. National Research Council. 1977. Leucaena: promising forage and tree crop for the tropics. Washington, D.C.: National Academy of Sciences. .1979. Tropical legumes: resources for the future. Washington, D.C.: National Academy of Sciences. .1980. Firewood crops: shrubs and trees for energy production. Washington, D.C.: National Academy of Sciences. .1982. Priorities in biotechnology research for international development. Washington, D.C.: National Academy of Sciencies. .1983. Mangium and other fast-growing acacias for the humid tropics. Washington, D.C.: National Academy of Sciences. Nitrogen Fixing Tree Association (NFTA) 1986. Erythrinas provide beauty and more. NFT Highlights 86-02. O'Connell, A.M., T.S. Grove and N. Malajczuk. 1979. Nitrogen fixation in the litter layer of eucalypt forests. Soil Biol. Biochem. 11: 681-2. Okigbo, B.N. 1984. Nitrogen-fixing trees in Africa: priorities and research agenda in multiuse exploitation of plant resources. Pesq. Agropec. Bras. 19: 325-30. Orchard, E.R. and G.D Darby. 1956. Fertility changes under continued wattle culture with special reference to nitrogen fixation and base status of the soil. In Comptes Rendus du Sixieme Congres. Inter. Science du Sol, Paris. Poschen, P. 1986. An evaluation of the Acacia albida-based agroforestry practices in the Hararghe highlands of Eastern Ethiopia. Agroforestry Systems 4: 129-43. Puppo, A., L. Dimitrijevic, H.G. Diem and Y.R. Dommergues. 1985. Homogeneity of superoxide dismutase patterns in Frankia strains from Casuarinaceae. FEMS Microbiol. Lett. 30: 43-6. Reddell, P.W. 1986. Management of nitrogen fixation by Casuarina. ACIAR Forestry Newsletter 2 (Sept./Oct): 1-3. Reddell, P. W., G.D. Bowen and A.D. Robson. 1986. Nodulation of Casuarinaceae in relation to host species and soil properties. Aust. J. Hot 34: 435-44. Roskoski, J.P. 1981. Nodulation and N2 fixation by Inga jinicuil, a woody legume in coffee plantations. I. Measurements of nodule biomass and field C2H2 reduction rates. Plant Soil 59: 201-6. .1982. Nitrogen fixation in a Mexican coffee plantation. Plant Soil 67: 282-292. .1986. Future directions in biological nitrogen fixation research. In Biotechnology of nitrogen fixation in the tropics (BIOnifT), Proceedings UNESCO Regional Symposium and Workshop, UPM, Malaysia, 25-29 August 1986 (in press). Roskoski, J.P., J. Montano, C. Van Kessel and G. Castilleja. 1982. Nitrogen fixation by tropical woody legumes: potential source of soil enrichment. In P.H. Graham (ed.), Biological nitrogen fixation technology for tropical agriculture. Cali, Colombia: CIAT. Sanchez, P.A. and J.G. Salinas. 1981. Low-input technology for managing oxisols and ultisols in tropical America. Adv. Agron. 34: 279-406. Sanginga, N., K. Mulongoy and A. Ayanaba. 1985. Effect of inoculation and mineral nutrients on nodulation and growth of Leucaena leucocephala. In H. Ssali and S.O. Keya (eds.), Biological nitrogen fixation in Africa. Nairobi: MIRCEN. .1986. Inoculation of Leucaena leucocephala (Lam.) de Wit with Rhizobium and its nitrogen contribution to a subsequent maize crop. Biological Agriculture and Horticulture 3:347-352. Schenck, N.C. 1982. Methods and principles of mycorrhizal research. Saint-Paul, Minnesota:American Phytopathological Society. Silvester, W.B. 1977. Dinitrogen fixation by plant associations excluding legumes. In R. W.F. Hardy and A.H. Gibson (eds.), A treatise on dinitrogen fixation. New York: John Wiley. .1983. Analysis of nitrogen fixation. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems: foundations and applications. The Hague:Nijhoff/Junk. Somasegaran, P. and HJ. Hoben. 1985. Methods in legume-Rhizobium technology. University of Hawaii NifTAL Project and MIRCEN, Hawaii. Sougoufara, B., E. Duhoux and Y.R. Dommergues. 1987. Improvement of nitrogen fixation by Casuarina equisetifolia through clonal selection. Arid Soil Research and Rehabilitation 1:129-132. Sprent, J. 1983. Agricultural and horticultural systems: implications for forestry. In J.C. Gordon, C.T. Wheeler and D.A. Perry (eds.), Symbiotic nitrogen fixation in the management of temperate forests. Corvallis, Oregon: Forest Research Laboratory. Sumberg, J.E. 1985. Collection and initial evaluation of Gliricidia sepium from Costa Rica. Agroforestry Systems 3: 357-61. Tarrant, R.F. 1983. Nitrogen fixation in North American forestry: research and application. In J.C. Gordon and C.T. Wheeler (eds.), Biological nitrogen fixation in forest ecosystems: foundations and applications. The Hague: Nijhoff/Junk. Taylor, D.P. 1976. Plant nematology problems in tropical Africa. Helminthological Abstracts (Ser. B) 45: 269-84. Thiagalingam, K. 1983. Role of Casuarina in agroforestry. In S.J. Midgley, J. W. Turnbull and R. D Johnson (eds.), Casuarina ecology, management and utilization. Melbourne: CSIRQ. Torrey, J. and J. D. Tjepkema. 1979. Preface and program. Bot. Gaz. 140 (suppl), i-ii. Turnbull, J.W., D.J. Skelton, M. Subagyono and E.B. Hardiyanto. 1984. Seed collecting tropical Acacias in Indonesia, Papua New Guinea and Australia. Forest Genetic Resources Information (FAO, Rome) 12: 2-15. Virginia, R.A., W.M. Jarrell, D.H. Kohl and G.B. Shearer. 1981. Symbiotic nitrogen fixation in Prosopis (Leguminosae) dominated desert ecosystems. In A.H. Gibson and W.E. Newton (eds.), Current perspectives in nitrogen fixation. Canberra: Australian Academy of Science, von Carlowitz, P.G. 1986. Multipurpose tree and shrub directory. Nairobi: ICRAF. Walsh, K.B., B.H. Ng and G.E. Chandler. 1984. Effects of nitrogen nutrition on xylem sap composition of Casuarinaceae. Plant Soil SI: 291-3. Waring, H.D. 1985. Chemical fertilization and its economic aspects. In J. Burley and J.L. Stewart (eds.), Increasing productivity of multipurpose species. Vienna: IUFRO. Wiersum, K.F. 1985. Acacia meamsii. Multipurpose highland legume tree. NFT Highlights 85-02. Williams, P.K. 1984. Current use of legume inoculant technology. In M. Alexander (ed.), Biological nitrogen fixation: ecology, technology and physiology. New York: Plenum Press. Wilson, G.F., B.T. Kang and K. Mulongoy. 1986. Alley cropping: trees as sources of green manure and mulch in the tropics. Biological Agriculture and Horticulture 3: 251-67. Yadav, J.S.P. 1983. Soil limitations for successful establishment and growth of Casuarina plantations. In S.J. Midgley, J.W. Turnbull and R.D. Johnston (eds.), Casuarina ecology, management and utilization. Melbourne: CSIRO. Zhang, Z. and J. Torrey. 1985. Biological and cultural characteristics of effective Frankia strain FPCcl3 (Actinomycetale) from Casuarina cunninghamiana (Casuarinaceae). Annals of Botany 56: 367-78. Zhang, Z., M.F. Lopez and J.G. Torrey. 1984. A comparison of cultural characteristics and infectivity of Frankia isolates from root nodules of Casuarina species. Plant Soil 78:79-90.
1 In the field of forestry the term provenance refers to natural populations of trees originating in a specific geographic location (also see Burley, the volume). 2 Basamid, BASF, 140 rue Jules Guesde, BP 87,92203 Levallois-Perret Cedex, France. 3 NifTAL - Nitrogen Fixation by Tropical Agricultural Legumes, University of Hawaii, 14000 Holomua Road, Paia, Maui, HI 96779, USA. |