An e-publication by the World Agroforestry Centre
AGROFORESTRY A DECADE OF DEVELOPMENT 

|
|
An e-publication by the World Agroforestry Centre |
|
AGROFORESTRY A DECADE OF DEVELOPMENT |
|
|
section 3 Chapter 9 Henry N. Le Houérou Introduction Most of the natural grazing lands in intertropical Africa, i.e., those under summer rainfall regimes, can be called "silvopastoral", for most are found in either dry forest, woodland, bushland, wooded savanna or shrubland.1 Purely herbaceous grazing lands, i.e., grass savanna or "grassland" sensu stricto, are rather rare and restricted to particular conditions of soil, climate and/ or management: for instance on Vertisols, some types of shallow soils, or wet soils, or under very arid climatic conditions such as in the northern Sahel, or some southern African velds, or under management practices that tend to eliminate woody species. In Africa north of the Tropic of Cancer (Mediterranean Africa), under climatic conditions characterized by a winter rainy season, silvopastoral systems of livestock production are more or less confined to semi-arid to perhumid bioclimates. The North African arid zone is, to a very large extent, dominated by a dwarf-shrub type of steppe vegetation and therefore the livestock production systems developed in such grazing lands cannot be called silvopastoral. The African desert systems, characterized by contracted types of vegetation clustered along the drainage network, can, perhaps somewhat paradoxically, be called silvopastoral systems since they include both shrubs and trees, though these are very sparse. These different silvopastoral systems represent about 80 percent of the African natural grazing lands (Le Houérou, 1977). Shrubs and trees in the silvopastoral production systems constitute the basic feed resource of more than 500 out of the 660 million head of livestock (FAO, 1985), i.e., 165 and 218 million tropical livestock units (TLU)2 respectively. A number of studies suggest that ligneous species represent an average 10-20 percent of the overall annual stock diet in these production systems in terms of dry-matter uptake, but much more in qualitative terms as one of the main sources of protein and minerals in the diet, particularly during the dry seasons. In addition to their fodder value, trees and shrubs (sometimes collectively referred to in agroforestry jargon as "trubs") play an essential role, not only as fodder reserve for critical periods, but also as a multipurpose resource, fulfilling many basic needs of these pastoral populations. For example, they are a source of energy and fuel, fibres for clothing and handicrafts, wood for construction, tools, human food, medicines for man and animals, drugs and dyes, shelter for man and beast (zribas, bomas), maintenance of soil fertility via nutrient cycling, soil protection from erosion agents, etc. In various revegetation projects, such as reforestation, establishment of forage reserves, watershed management, sand-dune stabilization and agroforestry, the question often arises: what species to select; exotics or native? Common sense would suggest avoiding any dogmatism in such matters, and the taking of a pragmatic view based on experience. But this does not always happen: advantages of both exotics and native species are advanced. This, in my judgement, is a false dilemma. One should select whatever has proved best adapted and most productive under the circumstances and for the objective that is being pursued without undue consideration to ideological principles! In some instances exotics may prove best suited while in other circumstances native species may be preferable for various reasons such as self-regeneration, the wishes of local users, etc.
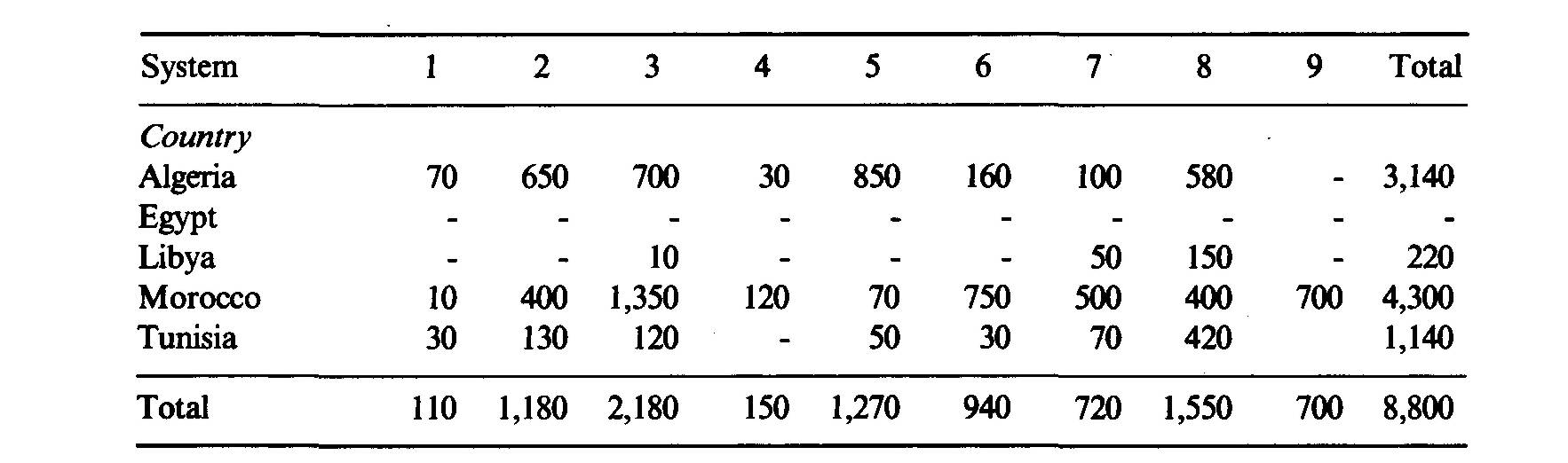
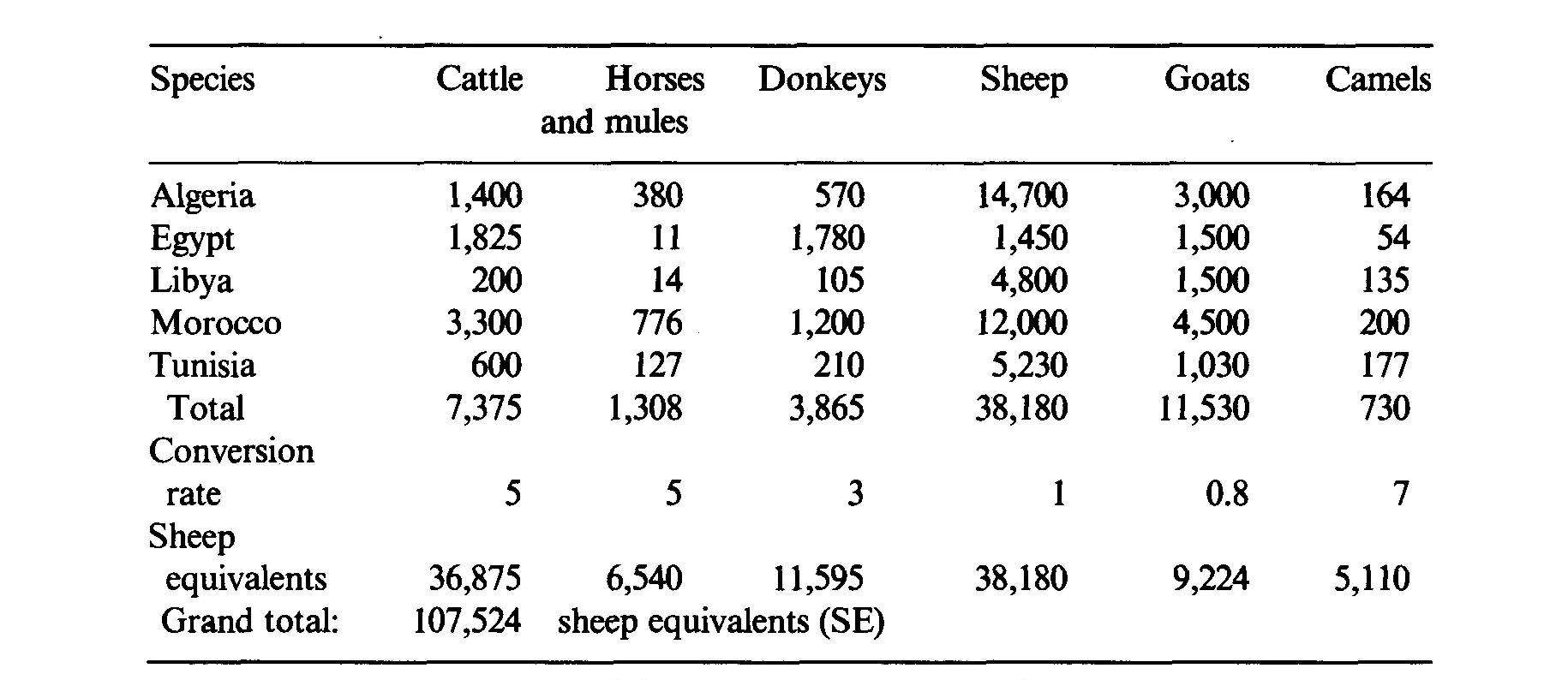
Africa north of the Tropic of Capricorn, which is considered in this paper, is 96 percent of the continent's surface area, that is, 28.2 million km2 including the islands. This area is extremely varied in terms of climate: Mediterranean, tropical, equatorial, subtropical, sub-equatorial, montane, and Afro-Alpine climates are found (Le Houérou and Popov, 1981). Most of the soil classes of the world are represented to a greater or lesser extent (FAO, 1974, 1978). The African flora comprise some 36,000 species of flowering plants, 30,000 of which belong to the intertropical zone (Brenan, 1979). About 9,000 of those are trees or shrubs (Wickens, 1980). About 75 percent of the latter (6,750) are browsed (Whyte, 1974), and over 4,500 have been recorded as being useful to man (Wickens, 1980). Shrubland, woodland and savanna communities occupy around 10,000 km2, i.e., 35 percent of the continent, the remainder being desert (36 percent), dense forest (22 percent) and cropland (7 percent) (Le Houérou, 1977). Mediterranean Africa Silvopastoral systems in the Mediterranean region of Africa are different in their biological composition, structure and physiognomy from those in intertropical systems. However, some similarities may be found with the highland silvopastoral systems of East Africa, with quite a number of common genera such as Pistacia, Olea, Rhus, Osyris, Jasminum, Colutea, Buxus, Ceratonia, Asparagus, Juniperus, Argyrolobium, and a physiognomy and structure that are strongly reminiscent of the Mediterranean garrigue and maquis with sclerophyllous shrubs and forests with acicular-leaved trees (Juniperus, Podocarpus) (Le Houérou, 1984). The area of the main types of silvopastoral systems of the Maghrib countries is shown in Table 1. The table shows that there is a total of about 8.8 million hectares of silvopastoral systems in northern Africa. These silvopastoral systems contribute substantially to the nutrition of some 10 million head of sheep-equivalents in the region (Table 2). The silvopastoral systems are estimated to provide about 10 percent of the total feed consumption by the regional livestock population, the balance coming from dwarf-shrub steppes, stubble and fallows, fodder crops, crop residues, agro-industrial by-products and concentrated feed, including cereal grain (mostly barley). The annual productivity of these silvopastoral systems in the region is in the neighbourhood of 400 Scandinavian feed units (SFU)3 per hectare (Le Houérou, 1976b, 1980b). Productivity varies widely from one system to another, with values ranging from 100 to 1,000 SFU ha-1yr-1. Similarly, the period of utilization varies greatly between systems; the highland systems (Juniperus thurifera, Abies sp., Cedrus atlantica) are mainly used as summer pastures within transhumant systems of animal husbandry. The lowland systems are used differently by various livestock species: cattle and sheep use them when stubble and fallow are not available from late fall to early summer, whilst goats use them all the year round. The transhumant and nomadic systems of northern Africa are on the decline, except for very short seasonal transhumance of a few kilometres. Animal production in these silvopastoral systems represents 60-80 percent of their productivity in economic terms (Le Houérou, 1976b, 1980a), the balance being fuelwood, charcoal, honey, distillation of essential oils, cork and some timber. These North African silvopastoral systems have been the subject of extensive study and review by the author (Le Houérou, 1973, 1976a, 1978, 1980a,b,c, 1981a, b, 1987a, b, c, d). The forest and woodland, and therefore the silvopastoral systems, are on the decline in northern Africa. This decline has two aspects: the reduction of the total area and a continuous decline of productivity per unit area. . The reduction of area under wooded land (forest, woodland and shrubland) is estimated to be of the order of 1 percent per annum (Le Houérou, 1981 a; Bourbouze, 1982). There are two reasons for this:
System 1: Quercus faginea forest and woodland; humid and perhumid zone. System 2: Quercus suber forest and woodland; subhumid and humid zones. System 3: Quercus ilex and Q. coccifera shrublands and woodlands; semi-arid and subhumid zones. System 4: Cedrus atlantica forest and woodland; cold subhumid and humid zones. System 5: Pinus halepensis forest and woodland, semi-arid to subhumid zones. System 6: Tetraclinis woodlands and shrublands; arid and semi-arid zones. System 7: Olea europaea sylvestris and Pistacia lentiscus shrublands; semi-arid to humid zones. System 8: Juniperus phenicea shrublands; arid and semi-arid zones. System 9: Argania sideroxylon parkland and shrubland; arid and desert zones of S. W. Morocco. Lesser areas under other systems include: System 10: Pinus pinaster mesogeensis, high rainfall zones, acidic soils hi Tunisia (2,000 ha), Algeria (1,200 ha), Morocco (15,000 ha). System 11: Abies moroccana, in high elevation: 5,500 ha in N. Morocco; Abies numidica, in high elevations: 1,000 ha in N. Algeria System 12: Juniperus thurifera, at elevations above 2,500 m: 31,000 ha hi Morocco and 1,000 ha in Algeria. System 13: Cupressus atlantica: 9,500 ha hi Morocco; Cupressus sempervirens: 100 ha in Algeria and 100 ha in Tunisia. System 14: Quercus toza (deciduous oak): 14,000 ha in N. Morocco. System 15: Pinus nigra: 100 ha hi N. Morocco. System 16: Cupressus dupreziana, a few hectares hi a deep wadi bed hi the Tassili n'Ajjerhi the central Sahara of Algeria.
Table 2 Livestock population in northern Africa (103 adult head)
Annual feed requirement: 108 million SE x 300 SFU =
32.4 billion SFU. Production from silvopastoral systems = 3.0 billion
SFU (from about 9 million ha). The protection of forest by state agents is a matter of:
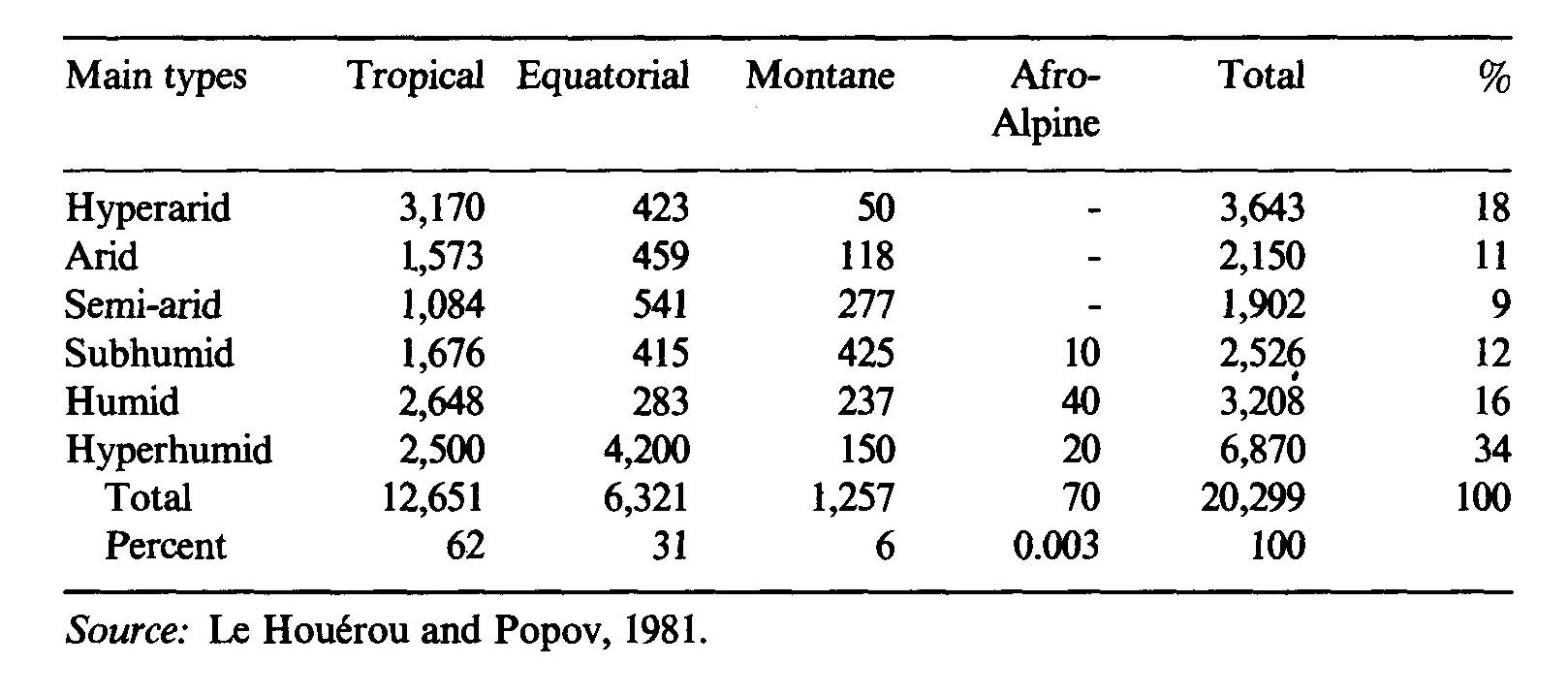
The decline in productivity is a result of over-exploitation — overstocking, overgrazing, overbrowsing, excessive woodcutting. Trampling, runoff, erosion, loss of soil fertility and loss of productivity of the overall ecosystem ensue. The current level of productivity is often only a third of that in reasonably well managed ecosystems and as low as a tenth in extreme cases: rain-use efficiency (RUE) factors of 0.5-1.0 kg DM ha-1 yr-1 mm-1, instead of the usual 4-6, are common (Le Houérou, 1984). Intertropical Africa General climate Intertropical Africa is a large area of some 22 million km2 encompassing a number of ecoclimatic zones (Le Houérou and Popov, 1981). Precipitation occurs mostly during the long-days season (summer), although there are monomodal and bimodal rainfall patterns. The number of rainy seasons depends usually on latitude: one rainy season in the 10-23° latitudes N and S and two seasons in the 0-10° latitudes on both sides of the equator. The monomodal regime is called "tropical" and the bimodal "equatorial". Both tropical and equatorial intertropical climates have dry (desert) and wet (rain forest) subtypes, which tend to become "amodal" (no rains or continuous rains) in extreme situations. Frost may occur in higher elevations. The lower altitude of frost occurrence naturally depends on the latitude: about 1,000 m a.s.l. near the Tropics of Cancer and Capricorn and around 2,500 m a.s.l. at the equator — that is, an increase of about 63 m for each degree of decrease in latitude or 0.57 m per km towards the equator. The isoline of year-long night frost is about 1,500 m higher, i.e., 2,500 m at the Tropics of Cancer and Capricorn and 4,000 m at the equator, and the lapse rate with latitude is similar. Frost occurrence gives rise to subtropical and subequatorial climates according to the number of rainy seasons. These montane climates in Africa are ecologically quite peculiar: they are mainly located in eastern and southern Africa. The areas with year-long night frost are referred to as Afro-Alpine: they too have very peculiar types of vegetation. The areas occupied by the main ecoclimatic zones in intertropical Africa are shown in Table3.
Desert silvopastoral systems In the desert zone, silvopastoral systems are restricted to the drainage network in or along wadis which originate in montane areas such as the Ahaggar, Tibesti Air and Adrar. These are the only sites used by desert pastoralists. Because of episodic runoff and deep underflows, permanent vegetation is present with more-or-less scattered individual trees and shrubs such as Acacia tortilis var. raddiana, A. ehrenbergiana, Balanites aegyptiaca, Maerua crassifolia, Capparis decidua, Grewia tenax, Salvadora persica, Ziziphus lotus saharae, Leptadeniapyrotechnica, Ochradenus baccatus, Aervapersica, Tamarixaphylla, Moringa peregrina, Nucularia perrini, Traganum nudatum, Cornulaca monocantha. These are mixed with herbs, legumes and perennial grasses such as Panicum turgidum, Lasiurus hirsutus, Cymbopogon proximus, Pennisetum dichotomum, Stipagrostis pungens and a palatable perennial desert sedge, Cyperus conglomerate. East African deserts have a similarly large number of tree and shrub species in the silvopastoral vegetation that colonizes the wadis and depressions of the Danakil, Somali and Chalbi deserts: Acacia raddiana, A. tortilis tortilis, A. t. spirocarpa, A. nubica, A. edgeworthii, A. reficiens, Balanites aegyptiaca, Hyphaene thebaica, Salvadora persica, Boscia sp., Cadaba sp., Dobera glabra mixed with dwarf shrubs such as Duospermum eremophilum, Indigofera spinosa, and Sericocomopsis hildebrandtii. There are also perennial grasses such as Panicum turgidum, Chrysopogon plumulosus, Cenchrus ciliaris, Andropogon spp. and a number of annual grasses of low value such as Aristida adscensionis, A. mutabilis, Chloris pycnothrix, and Tetrapogon cenchriformis. These systems are used in the same way as in the Sahara by nomadic pastoral tribes such as the Afar, Issa, Somali, Boran, Rendille and Turkana. These silvopastoral zones in the deserts are the very basis of these pastoral systems since there is virtually no perennial vegetation outside the silvopastoral depressions and wadis. More than any other, these desert silvopastoral systems are threatened by desertisation for the same reason of demographic growth as in northern Africa. But without these systems life cannot be sustained in the desert zones. Arid and semi-arid silvopastoral systems These systems cover large areas in the Sahel (3 million sq km), in East Africa (Ogaden, Somalia, northern Kenya, northern Tanzania), and Botswana, Kalahari and Namibia in southern Africa (nearly 4 million sq km). Arid and semi-arid silvopastoral systems cover about a third of intertropical Africa and sustain about 60 percent of the livestock population of the continent (Le Houérou, 1977). These silvopastoral systems are essentially savannas with various densities of shrubs and trees from less than 100 to over 3,000 individual "trubs" per hectare, dotting a herbaceous layer of annual grasses and herbs. The canopy area of the ligneous stratum also varies greatly from less than 1 percent to over 30 percent. The arid zone (100-400 mm of annual rainfall) is dominated by species in the genus Acacia and related genera of the Mimosoideae subfamily of legumes. In East Africa, species in the genus Commiphora are often found with the acacias. The semi-arid zone (400-600 mm of annual rainfall) is more varied in terms of woody cover. The Combretaceae often play a major role (Combretum spp., Terminalia spp., Anogeissus sp., Guiera sp.), but there are still a few acacias, particularly A. seyal, on fine-textured soils, but also A. Senegal, A. mellifera in the Sahel, A. etbaica, A. drepanolobium and A. btissei in East Africa, whilst agrosilvopastoral systems are dominated by Acacia albida (Faidherbia albida). There are about 100 main species of browse in the silvopastoral systems of the Sahel and about as many in the arid and semi-arid zones of eastern and southern Africa. About 30 percent of the species are common to the three regions (Walker, 1980; Lamprey et al, 1980; Le Houérou, 1978,1980a,b; von Maydell, 1983, this volume). In terms of animal nutrition woody species play a major role in these systems because they are the only source of protein, carotene and phosphorus during the long dry season and therefore livestock and wildlife cannot survive without them. This fact was not fully realized by scientists until recently, but it is now well accepted among livestock scientists, wildlife specialists, ecologists and range scientists (McKell, et al, 1972; McKell, 1987; Le Houérou, 1973, 1980a, b). Among the woody species, Capparidaceae play a major role in arid zones because of their high content of nitrogen and minerals (15-25 percent protein and 8-16 percent of non-silica minerals in the dry matter). Particularly significant genera in this respect are Maerua, Cadaba, Boscia, Crataeva, and Capparis (Lamprey et al, 1980; Le Houérou, 1980c). In the present socio-economic context, the role of browse in silvopastoral systems is to provide stability and productivity of livestock production, which is the major source of livelihood and income in arid and semi-arid African zones. The role they play could, in principle, be substituted by various concentrated feeds such as non-protein nitrogen (urea), salt licks and molasses, as practised in climatically comparable situations in developed countries such as the USA and Australia, but, given the terms of trade between livestock products on the one hand and the cost of transport and industrial products and by-products on the other, solutions of this type will not be used in pastoral zones of intertropical Africa in the foreseeable future. Silvopastoral systems in the subhumid and humid climates Subhumid and humid silvopastoral systems cover very large areas in Africa: most of the Sudanian ecological zone from the Atlantic Ocean to Lake Victoria and the Indian Ocean, and large parts of eastern and southern Africa, including the miombo, with a total area of 6-7 million sq km, i.e., about a third of intertropical Africa. Some 20 percent of the African livestock population and a large, but unknown, proportion of its wild ungulate populations live in this area. The livestock population amounts to 120-130 million head or 40-50 million TLU. These are wooded savannas of the Sudanian ecological region, and the homologous zone in the southern hemisphere is the miombo. It also includes the so-called derived savannas of the Guinea-Congolese rain forest ecological zone. The structure and function of this type of ecosystem are quite different from those of the arid and semi-arid zone, for various reasons. First, there are three very distinct sets of components: trees, shrubs and tall grasses. Capparidaceae and Mimosoideae do not play the major role they do in drier climates: the botanical composition of the woody layers is more diversified, with the exception of the miombo. Trees of the Leguminosae family, mainly Papilionoideae and Caesalpinioideae, are actually dominant in the miombo. Perennial tall grasses are dominant in the herbaceous layer — these include many Andropogonoideae, particularly the genera Andropogon, Hyparrhenia and related genera (Le Houérou, 1977). Secondly, bush fires play an essential role in the ecosystem, particularly as regards the grazing resource, since they induce regrowth of the perennial grasses in the dry season, thus easing the feed shortage at that period. Thirdly, soils are very different: large areas have ferruginous tropical soils and ferralitic hardpans, which practically prohibit cultivation on the interfluves. Fourthly, climate is more favourable with a rainy season lasting 5-9 months in the Sudanian and miombo zones and 10 or more months in the Guinea-Congo zone. Finally, livestock husbandry is impaired by trypanosomiasis since most of these silvopastoral zones are infested to a greater or lesser degree by tsetse flies (Glossina spp.), the vectors of the trypanosomes. As a consequence, animal husbandry is not of great importance in these zones as long as they remain silvopastoral in nature since the tsetse flies only live in areas with some tree or bush cover. Indeed, one of the methods of elimination of trypanosomiasis is destruction of the woody layer, hence of the silvopastoral system! However, there are trypanotolerant breeds of stock, such as, for cattle, the NTJama breed originating in the mountains of Fouta Djalon and Nimba in Guinea and the West African shorthorns, a complex of breeds from Ghana, Nigeria and Ivory Coast. Smallstock also have trypanotolerant breeds such as the Djalonke sheep of Guinea and neighbouring countries, and the achondroplasic West African dwarf goat of Nigeria, Ghana, Ivory Coast and neighbouring countries. These breeds lend themselves to some livestock husbandry in tsetse-infested areas. Some countries, such as Ivory Coast and Zaire, have long since initiated a policy of meat production based on these trypanotolerant breeds, particularly the N'Dama, rather than the "negative" policy adopted in other countries of destroying the tree layer in order to permit an animal-husbandry system based on exotic breeds, possibly in conjunction with the intensive use of chemicals. The policy of encouraging local trypanotolerant breeds seems a sound one because various tsetse-eradication schemes have rather poor records and trypanotolerant breeds allow the preservation of ecological diversity, including maintaining a tree layer. In the humid and sub-humid climates, the silvopastoral systems do not play the major role that they do in arid and semi-arid situations. This is particularly so because there are viable alternatives to the use of browse in the relatively shorter dry seasons in these regions. For example, the most currently used alternative is burning. The burning of the tall Andropogon grass at the beginning of the dry season allows some regrowth a few weeks later which provides a small amount of fodder of high quality that would adequately complement the roughage available in the unburnt areas. A skilful use and timing of controlled burning may thus ensure an adequate diet for stock throughout the dry season. This method has been used for decades, particularly in large meat-production ranches based on the N'Dama breed at Mwesi in southern Zaire (Le Houérou, 1977). Nevertheless, in areas of mild tsetse infestation browse remains a major component of livestock diet during the dry season in the subhumid zones, especially between the isohytes of 600 and 1,200 mm of mean annual rainfall where both Zebu cattle and Taurine trypanotolerant cattle co-exist and interbreed (e.g., the West African Mere, the Sanga and the Nilotic Ankole). This remark is also valid for the dwarf goat in the humid zones of West Africa which consumes a large proportion of browse in its diet and plays an important role in the household economy of the small West African farmers (Mecha and Adegbola, 1980). Highland (montane) silvopastoral systems Montane silvopastoral systems do not represent very large areas in Africa, but they harbour a large livestock population. Together with the Afro-Alpine ecological zone, they cover some 50,000 sq km and support 120-150 million head (almost 50 million head for Ethiopia alone), or 20-30 percent of the intertropical African livestock population. These montane silvopastoral systems play an essential role, particularly during the rainy season, unlike the systems in arid and semi-arid zones. This is because these montane areas are intensively cultivated so that stock have nowhere to go during the cropping season except to silvopastoral areas on rugged terrain. During the dry post-harvest season the stock mostly feed on crop residues. Moreover, the montane silvopastoral systems are also used as transhumant grazing land for neighbouring pastoral arid zones during dry seasons. Thus these areas are used by resident herds in the rainy season and by transhumant pastoralists during the dry season. They are thus heavily exploited all the year round, and, as a result, they are heavily depleted in terms of grazing value in spite of the favourable climatic conditions. They are therefore covered by herbaceous species of little value such as Pennisetum schimperi, P. villosum, and Eleusine jaegeri, the shrub and tree layers having been heavily chopped, lopped, overbrowsed, cut down, or eradicated. The role of shrubs and trees in these montane silvopastoral systems is difficult to quantify or even evaluate. The only thing which is certain is that they are receding at a rapid pace. A study on a number of test zones in the Ethiopian highlands, based on the comparison of aerial photos taken 20 to 25 years apart, yielded frightening results (Haywood, personal communication).
As mentioned above, the shrubs and trees in the silvopastoral systems of Africa have a multiplicity of roles, and the literature on them is extensive. Therefore the topic will be dealt with in very general terms with special emphasis on two subjects, namely, browse and the screen function. Traditional uses There are innumerable traditional uses of shrubs and trees. Some African languages, for instance, use the same word to mean tree and medicine (Dia, 1986). The traditional uses may be grouped into 13 main headings:
Generally speaking almost all parts of trees are used in one way or another. To cite one example, Acocanthera schimperi in the East African montane zone produces delicious fruits for human consumption, is a good firewood species, and the bark of the roots is used, after some preparation, to make a lethal arrow poison! Browse The browse aspect of trees and shrubs has been reviewed in some detail by the author (Le Houérou, 1980a) for western and southern Africa, by Lamprey et al. (1980) for East Africa, by Walker (1980) for southern Africa, and by Lawton (1980) for the miombo. There are about 200 main browse species in intertropical Africa. Special mention needs to be made of legumes (mainly Mimosoideae and Caesalpiniodeae), Capparidaceae, Combretaceae, Tiliaceae, Rubiaceae and Rhamnaceae, in decreasing order of importance, in animal diets. Average values of chemical composition and feed value, based on a review by the author (Le Houérou, 1980c) for some 850 samples and 150 species from East and West Africa are: crude protein 13 percent of the DM, crude fibre 24 percent, fat 4 percent, nitrogen-free extract 48 percent, ash 11 percent, phosphorus 0.18 percent and calcium 1.75 percent. Capparidaceae yield an average crude protein content of 20 percent, while legumes average 16 percent. Leaves of some Capparidaceae have an average crude protein content of 30 percent, which makes them similar in this respect to protein-rich concentrated feed! A figure of 44 percent crude protein has been reported in the leaves of Justicia salviojdes a species of Acanthaceae from the miombo (Lawton, 1980). Feed value has been estimated using various in vivo trials on digestibility and animal performance. For leaves and shoots, DM digestibility is, in general, 50 ± 5 percent. There are exceptions of very low digestibility (25-35 percent) as well as very high figures of 60-70 percent in vivo (Mabey and Rose-Innes, 1964, 1966). Some specialists of animal production have questioned the validity of the proximal analyses data, as well as those concerning digestibility of energy or protein, due to the presence of tannins and other polyphenolic compounds that may inhibit the actual assimilation of nitrogen and sugars, particularly in the intestine. The author's studies on animal performance over long periods, using sheep feeding only on known quantities of selected browse, do not confirm the negative impact of browse on sheep performance. On the contrary, the experiments carried out continuously over nearly one year, showed that animals feeding on the 10 species of browse under experiment showed a performance in direct relation to the amount of feed consumed. Moreover, sheep feeding partly on range and receiving a browse complement in the pen performed significantly better than those feeding on a pure range diet, even under excellent range conditions. The productivity of browse in arid and semi-arid silvopastoral systems was reviewed by the author (Le Houérou,-1980a). The order of magnitude of productivity is 1 kg DM ha-1 mm-1 of consumable biomass per year, where mm refers to average rainfall per year. That is about a third of the productivity of the herbaceous layer. This figure is, of course, liable to enormous variability depending on the ecosystem concerned and on its present and past management. The range is usually 0.1-5.0 kg DM ha-1 yr-1 mm-1. Production of 5,000kg DM ha-1 yr-1 has been reported from Acacia hockii in riverine conditions in East Africa (Pellew, 1980), a figure that is probably close to the maximum under natural conditions. Average individual "trub" production is of the order of 1 kg DM yr-1, with a range of 0.08-50.0 kg, most figures ranging from 0.5 to 5.0 kg (Bille, 1977; Poupon, 1980; Hiernaux, 1980; Dayton, 1978; Pellew, 1980; Kelly, 1973; Kennan, 1969; Rutherford, 1978; Cissé, 1980a, b, 1986). Equally important are the availability of browse in critical seasons and its direct accessibility to livestock and wildlife. The maximum standing crop of browse occurs naturally during the rainy season, but the most important is its availability during the dry season and the so-called "pre-rainy" season. In the Sahel, for instance, the most critical seasons are the hot dry season from March to May and the pre-rainy season in June-July. During these seasons, not more than a third of the maximum standing crop of that present in October is available to animals, although some of the dried leaves that fall are gathered by stock. Thus the amount of browse available at the most critical periods does not represent more than 20-25 percent of annual production. The maximum standing crop in September-October in the Sahel represents about 80 percent of annual production. But this does not tell the whole story because part of the crop can be stored in the form of dried pods such as those of Acacia albida, Acacia tortilis, Prosopis juliflora, and P. cineraria. These pods are traded in local markets for a monetary value close to that of similar concentrates of similar feed value, which also shows that local farmers and stock owners are well aware of the actual value of these products. The distribution and structure of available browse in space is another important issue. Smallstock can browse up to about 130 cm above the ground, cattle about 2 m, camels 3 to 4 m, and giraffe up to 5 m. The distribution of browse in these strata is thus an important point to consider in relation to the species of livestock which are kept (goats are known to be able to climb trees). Distribution of browse above the ground is extremely variable and depends on many factors: nature and botanical composition of the "trub" layer, kind of present and past management applied to it (trimming, lopping, etc.). In central Mali, for instance, Hiernaux and Cissé (1983) estimated that about 50 percent of the browse is located between the ground and a height of 2m, 30 percent between 2 and 3 m, and some 20 percent above 3 m. The horizontal distribution is also of relevance since the woody layer is usually not randomly distributed but follows a pattern more or less linked to runoff and drainage. This horizontal distribution, in turn, affects the nature of the species which are present and therefore affects livestock behaviour as well as the practices of the herder who will seek particular species in their preferred sites. In the Sahel, browse species consist of groups with specific characteristics which determine the type of management required (Gillet, 1986). Thus one could differentiate those species- having permanent or quasi-permanent green leaves such as Maerua crassifolia, Balanites aegyptiaca, Boscia senegalensis, B. agustifolia, B. salicifolia, Cadaba glandulosa and Salvadora persica (which all belong to the family Capparidaceae except Balanites and Salvadora). Then there are those species which leaf early at the end of the dry season (precession of leafing) such as Commiphora africana; these are used only for a short period in June-July before green grass becomes available. Commiphora africana is, therefore, a most important species at this particular time of the year, which is the most critical in the annual cycle when there is little feed available elsewhere and animals are emaciated by nine months of dry season and malnutrition. Moreover, females that are usually in a late stage of pregnancy at this time need protein-rich diets which only Commiphora can supply at that time. The winter flowering species also play a significant role as the flowers, shoots, young leaves and young fruits are consumed then. This is the case, for instance, with several Combretaceae (Combretum, Terminalia) and also with some of the Sudanian species such as Gardenia erubescens (Rubiaceae). Species which set leaves right at the onset of the rains, such as Combretum aculeatum and Feretia apodanthera, also need to be differentiated. These are also keenly sought species at a time of scarcity, and they play a role somewhat similar to Commiphora but in a somewhat different way. They are browsed throughout the rainy season and the first half of the dry season, whereas Commiphora is ignored as soon as green grass is available, and sheds its leaves soon after the last September rain. A fifth group is composed of species whose foliage is lopped and fed to penned or household animals, or prestige animals (household sheep — "moutons de case" — and horses). Among these, there are five main species in West Africa: Pterocarpus lucens, particularly in Mali and Niger, Pterocarpus erinaceus in southern Senegal, Khaya senegalensis in several countries, Terminalia avicennoides in Niger, and Acacia albida in many countries of Africa. A last group includes the species usually found around termitaria which remain green longer in the dry season than in the surrounding bush; they are therefore important browse species, particularly Grewia bicolor, Grewiavittosa, Boscia salicifolia, Maerua oblongifolia, Feretia apodanthera. The screen function The screen function is obvious from field observations: animals and people seek shade at midday when shade temperatures may reach 35-45° C. In depleted rangelands of the arid and semi-arid zone, grass-layer production is usually much higher (up to twice or more) under shade than in the open; and it remains green 4-6 weeks longer at the end of the rainy season. Measurements in the semi-arid zone of Botswana (25° S, 25° 50' E; 550 mm; 1,000 m altitude) over several years showed that under the canopy of Peltophorum africana, Acacia tortilis and Grewia flava, solar radiation and windspeed were reduced by about 50 percent as compared to a nearby open test area. As a consequence, potential evapotranspiration was reduced by 70 percent under the canopy, while the continuous grass layer of Panicum maximum had a production 26 percent higher when grazed and 12 percent when ungrazed (Pratchell, personal communication). Bille (1977) reported a production of the grass layer twice as large in the shade as compared to the open in the Sahel of Senegal (combined production of grass and shrubs). The photosynthetic efficiency during the growing season was 1.4 percent, whereas it barely reached 0.3 percent in the surrounding open grassland (global radiation 180 kcal cm-2 yr-1 (752 kJ); incident radiation 90 kcal cm-2 yr-1 (376.2 kJ); 1 g DM = 4.2 kcal cm-2 yr-1 (17.6 kJ); NPP = 125 percent of maximum standing crop). The efficiency of the system was thus 4.6 times greater in the multistorey vegetation structure as compared to a monostratum grass layer. These facts, however, should not be over generalized as there are instances where shrub cover may reduce production from grassland understorey, particularly when the ecosystem is in a good dynamic status (Le Houérou, 1984).
All monitoring studies in the Sahel and East Africa show a considerable regression of the woody cover over the past 30 years, caused by a combination of two phenomena: prolonged droughts from 1970 to 1985 and over-utilization resulting from the growth of human and stock populations which has been occurring at a rate of about 2.5 percent per annum. The comparison of aerial photographs taken at about 20-year intervals has given absolute proof of the substantial decline of the tree and shrub cover in Chad, Sudan, Niger, Mali, Burkina Faso, Senegal, Ethiopia, Somalia, and Kenya. The decline has been of the order of 20-35 percent between 1954 and 1975 and has accelerated since the early 1980s (Gaston, 1975; Haywood, 1981; De Wispelaere, 1980; De Wispelaere and Toutain, 1976; Gaston, 1981; Lamprey, 1975,1983; Lusigi, 1981; Boudet, 1972,1977; Le Houérou, 1977, 1980a, 198Ib; Peyre de Fabregues, 1985; Peyre de Fabregues and De Wispelaere, 1984; Gillet, 1986; Le Houérou and Gillet, 1986). These results are confirmed by field surveys over the whole Sahel; for instance the mean mortality of trees and shrubs during the 1969-1973 drought has been estimated at 40-50 percent. Over some areas the die-off was total. Detailed research on the functioning of the Sahelian ecosystems, conducted at Fete Ole in northern Senegal over a period of 10 years, including the 1969-1973 drought, showed an average mortality of 20 percent under conditions of total protection (Bille, 1977, 1978; Poupon, 1980). Naturally, under current conditions of overexploitation the situation is much worse, with mortalities often reaching 50 percent. A study of the ligneous population in Fete Ole showed that 30 years of total protection would be required to offset the effects of the 1969-1973 drought and to bring the production back to its pre-drought level (Bille, 1978). But the subsequent 1980-1985 period was far worse (Tucker et al, 1986; Le Houérou and Gillet, 1985; Peyre de Fabregues, 1985). According to most ecologists familiar with the Sahel situation, the Sahel might never recover from these two consecutive droughts (Peyre de Fabregues and De Wispelaere, 1986). The lack of regeneration after the droughts can be explained by three main factors:
Assuming that there will be three drought periods in the next hundred years — there have been four since the beginning of the present century — it is to be feared that the ligneous cover of the Sahel will be reduced to close to zero within the next 50 years (unless continuous regeneration could be ensured through good management). Such regeneration being virtually non-existent as environmental degradation worsens, the Sahel would become unexploitable by livestock outside the rainy season, unless animals are fed concentrates and/ or urea and minerals in order to offset the shortage of browse. Of course, such complementary feeding is possible in principle as it is used in other arid regions, though under totally different socio-economic conditions. However, this solution is not likely to be a viable one in the Sahel or under East African socio-economic conditions in the foreseeable future because of cost/benefit considerations and the lack of adequate managerial skills. Given current meat producer prices and prevailing offtake rates in pastoral production systems, scarcely any investment is economically feasible (though slight differences exist between countries in this respect). In fact, one cannot see any way in which livestock could be supplemented for 8-9 months of the year with the present prices of meat and concentrates (De Montgolfier-Kouevi and Le Houeiou, 1980; Le Houérou, 1987b). The destruction of the ligneous cover of browse species would therefore lead to a dead-end for the livestock industry in the Sahel and in large parts of East Africa before the middle of next century, if not earlier, if population growth continues unabated. Generally speaking, the situation is not yet as desperate in East Africa as it is in the Sahel; but locally it may be just as bad, particularly in Somalia, south-east and north-east Ethiopia and northern Kenya. An unmistakable indicator of environmental deterioration is when traditional cattle and sheep pastoralists shift to rearing camels and goats, that is from grazers to browsers. Some ethnic groups such as the Rendille and Samburu of northern Kenya have already made this shift.
As indicated previously, for nutritional and economic reasons, browse should represent 20 25 percent of domestic ruminants' intake (30 percent in the dry season, 5 percent in the rainy season) in the arid and semi-arid silvopastoral areas of Africa. How could such a balance be ensured? Or, to put it in another way, how could sound management be established on the principle of sustained maximum output? Such an objective would require a careful and co-ordinated course of action. At the conceptual and planning level, the philosophy and objectives ought to be clearly defined, and the strategy and means to attain the objective targets should be clearly spelt out at the outset and selected and described. This implies a dynamic livestock policy, including stratification of the industry, a marketing and a price policy giving a fair share to the producer, and also promotion of quality products. At the technical execution level, in the field, the problem is theoretically simple — if not easy. It is, after all, a question of adapting stocking rates to the carrying capacity of the ecosystem, that is an offtake of the primary production of not more than 25-30 percent. In practice this would mean that human and animal densities would be controlled and that simple techniques such as deferred grazing, periodic enclosures and the adaptation of watering regimes to the density and seasonal occurrence of watering points would be applied. Of course, this kind of management implies choices and daily decisions, i.e., the notion of responsibility. Yet the present situation in dry Africa is characterized by a general lack of responsibility at resource-management level, in other words in communal or common ownership of land and water. Water and pasture are actually common or public resources, whereas animals are privately owned, so it is in every user's interest to draw a maximum and immediate profit from the common resource without bothering about what may happen to it in the long run. Such a situation results in destruction of the common resources in the long run for the sake of the individual's own immediate benefit. This has been labelled the "looting strategy" (Le Houérou, 1977), or the "tragedy of the commons" (Hardin, 1968). It is quite obvious that no rational system of any kind can be implemented without the concept of responsible management, whether by individuals or groups. Meeting these responsibilities involves fundamental land reforms in terms of land tenure, and of land and water usufruct. Such systems, on a collective basis, existed long ago in pre-colonial Africa but with a much lower density of human and animal population so that they resulted in a fairly steady balance between the resources and their utilization. This equilibrium, which was certainly not ideal as regards productivity, human dignity or social justice, is badly endangered today by severe overutilization. If drastic socio-political reforms are not implemented without delay in order to ensure the rational management of shrubland ecosystems, the arid and semi-arid zones of Africa will have to face a very deep crisis threatening their present main resource. The situation in the subhumid and humid eco-climatic zones is certainly less gloomy, and development prospects are much better, at least on deep soils. But what will happen to the immense areas of shrublands and forests growing on shallow ferruginous or lateritic (ferralitic) hardpans? At present browse plays an important part in livestock production, especially during the second half of the dry seasons.
Bille, J.C. 1977. Etude de la production primaire nette d'un écosysteme sahelien. Trav. & Doc. de l'ORSTOM, No. 65, Paris. .1978. Woody forage species in the Sahel, their biology and use. Proceedings of the First International Rangeland Congress, Denver, Colorado. Boudet, G.G. 1972. Desertification de l'Afrique tropicale seche. Adansonia, ser. 2, 12 (4): 505-524. .1977. Desertification ou remontee biologique au Sahel? Cah. ORSTOM, Ser. Biol, XII, 4: 293-300. Bourbouze, A. 1982. L'elevage dans la montagne marocaine. Organisation de Pespace et utilisation des parcours par les eleveurs du Haul Atlas. Doct. Dissert. Inst. Nat. Agron., Paris-Grignon. .1986. Adaptation a diffeients milieux des systemes de production des paysans du haul Atlas. Techniques et Cultures 1, 59-94. Brenan, J.P. 1979. Some aspects of the phytogeography of tropical Africa. Ann. Missouri Bot. Garden 65:437-478. Cissé, M.I. 1980a. The forage production of some Sahelian shrubs: relations between maximum leaf biomass and various physical parameters. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA. .1980b- Effects of various stripping regimes on foliage production of some browse bushesof the Sudano-Sahelian zone. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA. .1986. Les parcours saheliens pluviaux du Mali Central. Caracteiistiques et principes echniques pour une amelioration de leur gestion dans le cadre des systemes de production animate existants. In LA. Toure, P.I. Dia and M. Maldague (eds.), Laproblematique et les strategies sylvo-pastorales au sahel. CIEM, Univ. de Laval, Quebec and UNESCO, Paris. Dayton, B.R. 1978. Standing crop of dominant Combretum species at three browsing levels in the Kruger National Park. Koedoe 21: 27-76. De Montgolfier-Kouevi, C. and H.N. Le Houérou. 1980. Study on the economic viability of browse plantations in Africa. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA. De Wispelaere, G. 1980. Les photographies aeriennes t6moins de la degradation du couvert ligneux dans un écosystème sahélien sénégalais. Influence de la proxirnit6 d*un forage. Cah.ORSTOM, sér. Sces Hum., XVII, 3-4: 155-166. De Wispelaere, G. and B.Toutain. 1976. Estimation de l'évolution du couvert végétal en vingt ans, conséutivement à la sécheresse dans le Sahel voltaïque. Rev. Photointerpr. 76: 3/2. Dia, P.I. 1986. Espace, ressources et pasteurs du sahel sénégalais. In LA. Touré, P.I. Dia and M. Maldague, L aproblématique et les stratégies sylvo-pastorales au sahel. CIEM, Univ. de Laval, Quebec and UNESCO, Paris. FAO. 1984. FAO Production Yearbook. Rome: FAO. .1985. FAO Production Yearbook. Rome: FAO. FAO/UNESCO. 1972/1975. World soil map 1/5,000,000: Africa. FAO, Rome and UNESCO, Paris. Gaston, A. 1975. Etude des pâturages du Kanem après la sècheresse de 1973. Farcha, Tchad: IEMVT. .1981. La végétation du Tchad: Evolutions r&entes sous influences climatiques et humaines. These Doct. Sees, University of Paris XII, IEMVT, Maisons-Alfort. Gillet, H. 1986. Principaux arbres fourragers du sahel senegalais. In LA. Touré, P.I. Dia and M. Maldague (eds.), La problematique et ks strategies sylvo-pastorales au sahel. CIEM, University of Laval, Quebec and UNESCO, Paris. Hardin, G. 1968. The tragedy of the commons. Science 162: 1243-1248. Haywood, M. 1981. Evolution de Putilisation des terres et de la v6getation dans la zone soudano-sahelienne du projet CIPEA au Mali. Addis Ababa: CIPEA/ ILCA. Hiernaux, P. 1980. Inventory of the browse potential of bushes, trees and shrubs in an area of the Sahel of Mali: methods and initial results. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA. Hiernaux, P. and M.I. Cissé.1983. Les ressources naturelles des systémes de production animate du Gourma. Etude exploratoire. Doc. de Progr. AZ/92 F, CIPEA, Bamako. Kelly, R.D. 1973. A comparative study of primary production under different kinds of land use in south eastern Rhodesia. Ph. D. dissertation, University of London. Kennan, T.C.D. 1969. The significance of bush in grazing land in Rhodesia. Rhod. Sclent. News 3: 331-336. Lamprey, H.F. 1975. Report on the desert encroachment reconnaissance in the northern Sudan. Nairobi: UNEP (mimeo). .1983. Pastoralism yesterday and today: the overgrazing problem. In F. Bourliere (ed.), Tropical savanna. Ecosystems of the world, Vol. 13. Amsterdam: Elsevier. Lamprey, H.F., D.J. Herlocker and C.R. Field. 1980. Report on the state of knowledge on browse in East Africa in 1980. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Lawton, R.M. 1980. Browse.in the Miombo. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Le Houérou, H.N. 1973. Ecologie, demographic et production agricole dans les pays m6diterraneensu tiers-monde. Options Mediterraneennes 17: 53-61. .1976a. Problemes et potentiality des terres arides nord-africaines. Options Mediterraneenes 26: 17-35. .1976b. The rangelands of North Africa: typology, yield, productivity and development. In Proceedings of the symposium on evaluation and mapping of tropical African rangelands. Addis Ababa: ILCA. .1978. The role of shrubs and trees in the management of natural grazing lands (with particular reference to protein production). Position paper, item No. 10, VIIIth World Forestry Congress, Jakarta, Indonesia and FAO, Rome. .1977. The grasslands of Africa: classification, production, evolution and development outlook. Proceedings of XIHth International Grassland Congress, Vol. I. Berlin, DDR: Akademie Verlag. .1980a. Browse in northern Africa. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA. .1980b. The role of browse in the sahelian and sudanian zones. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa, ILCA. .1980c. Chemical composition and nutritive value of browse in West Africa. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. .1981 a. The impact of man and his livestock on mediterranean vegetation. In F. di Castri, D.W. Goodall and R.L. Specht (eds.), Mediterranean-type shrublands. Ecosystems of the world, Vol. 11. Amsterdam: Elsevier. .1981b. Impact of the goat on Mediterranean ecosystems. Proceedings of the 32nd Annual Meeting of the European Association for Animal Production, Sheep and Goats Commission, Zagreb, Yugoslavia. .1984. Rain-use efficiency: a unifying concept in arid lands ecology. J. Arid Envir. 7:213-247. .1987a. The shrublands of Africa. In C.M. McKell (ed.), Shrub biology and utilization. N.Y.: Academic Press (in press). .1987b. An assessment of the economic feasibility of fodder shrub plantation in Africa. In C.M. McKell (ed.), Shrub biology and utilization. N.Y.: Academic Press (in press). .1987c. Inventory and monitoring of rangeland ecosystems in the Sahel. FAO, Rome and UNEP, Nairobi. .1987d. The grazing-land ecosystems of the Sahel. In Ecological studies. Heidelberg: Springer Verlag. Le Houérou, H.N. and H. Gillet. 1986. Conservation vs desertisation in the African arid lands. In M.E. Soulé (ed.), Conservation biology, the science of scarcity and diversity. Sunderland, Massachusetts: Sinauer Associates. Le Houérou, H.N. and G. F. Popov. 1981. An ecoclimatic classification of intertropical Africa. Plant roduction Paper No. 31, FAO, Rome. Lusigi, W.J. 1981. Combatting desertification and rehabilitating degraded production systems in Northern Kenya. Technical Report No. A-4. Nairobi: IPAL, UNESCO. Mabey, G.L. and R. Rose-Innes. 1964/66. Studies in browse plants in Ghana: Emp. J. of Exper. Agric.: XXXII, 126: 114-130; 127: 180-190; 128: 274-278; N. Ser., 2: 27-32, 113-117. McKell, C.M. (ed.). 1987. Shrub biology and utilization. New York: Academic Press. McKell, C.M., J.P. Blaisdell and R. J. Goodin (eds.). 1972. Wildland shrubs — their biology and use. USDA Forest Service, General Technical Report INT-I, Washington, D.C. Mecha, I. and T. A. Adegbola. 1980. Chemical composition of some southern Nigeria forage eaten by goats. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Pellew, R.A. 1980. The production and consumption of Acacia browse and its potential for animal protein production. In H.N. Le Houérou (ed.). Browse in Africa. Addis Ababa: ILCA. Peyre de Fabregues, B. 1985. Quel avenir pour l'elevage au Sahel? Rev, Elev. Medec. Vet. Pays Tropic. (38)4:500-508. Peyre de Fabregues, B. and G. De Wispelaere. 1984. Sahel: fin d'un monde pastoral? Marches Tropicaux 2488-2491 (12 Oct.). Piot, J. 1980. Management and utilization methods for ligneous forage: natural stands and plantations. In H.N. Le Houeiou (ed.), Browse in Africa. Addis Ababa: ILCA Addis Ababa. Piot, J. and I. Diaite. 1983. Systemes de production de'elevage au Senegal. Etude du couvert ligneux. CTFT, Nogent/ Mame and LNERV/ISRA, Dakar. Poupon, H. 1980. Structure et dynamique de la strate ligneuse d\me steppe sahélienne au Nord-Senegal. Trav. & Doc. de TORSTOM, No. 115, Paris. Rutherford, M.C. 1978. Primary production ecology in South Africa. In M.J.A. Werger, Biogeography and ecology of Southern Africa. The Hague: Junk. Tucker, C.J., C.O. Justice and S.D. Prince 1986. Monitoring the grasslands of the Sahel. J. of Remote Sensing 7 (11): 1571-1581. Tucker, C.J., C.L. Vanpraet, M.J. Sharman and G. Van Ittersum. 1985. Satellite remote sensing of total herbaceous biomass production in the senegalese Sahel: 1980-1984. Remote Sensing of the Environment 17: 233-249. von Maydell, H.-J. 1983. Arbres et arbustes du sahel. Eschborn, F.R.G: GTZ. Walker, B.H. 1980. A review of browse and its role in livestock production in Southern Africa In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Wickens, G.E. 1980. Alternative use of browse species. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Wilson, R.T. 1980. Fuelwood in a Central Malian town and its effects on browse availability. In H.N. Le Houérou (ed.), Browse in Africa. Addis Ababa: ILCA. Whyte, R.0.1974. The use and misuse of shrubs and trees as fodder: Africa. Joint publication No. 10, Imperial Agricultural Bureaux, Bureau of Pasture, Field Crops Forestry and Nutrition, Aberystwyth, Wales.
1 Woodland - open dry forest with 50% or
more canopy cover; 2 1 TLU = approx. 250 kg liveweight of animal. 3 1 SFU = 3 kg dry matter, being equivalent to tthe net energy of 1 kg barley. |