An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 4 Site Effects, Silviculture, and Symbiosis Session Papers Site Effects Variability in the Growth of Faidherbia a Termite Connection? J. Brouwer1, S.C. Geiger2, and R.J. Vandenbeldt Abstract
Introduction Spatial variability in the growth rate of crops and trees over distances of only 5-20 m is widespread in the Sahelian zone of West Africa. This effect is largely due to preexisting variability in soil chemical and physical properties (Scott-Wendt et al. 1989; Manu et al. 1990; Geiger and Manu 1991). The so-called 'albida effect', where soils under the canopy of Faidherbia albida trees have been found to be more fertile than soils in the adjacent open field is probably due to a considerable degree to such preexisting soil differences. Geiger et al. (in press) have shown that F. albida growth is particularly good around abandoned termite mounds, but that there were other areas of good growth as well. Improvement of soil fertility by termites is well-documented (Lal 1987, Lee and Wood 1971, Miedema and Van Vuuren 1977). Carrying on from that work, we investigated whether the other areas of good growth in the plantation might in fact be associated with higher levels of previous termite activity, n conditions but nevertheless and F. albida growth.
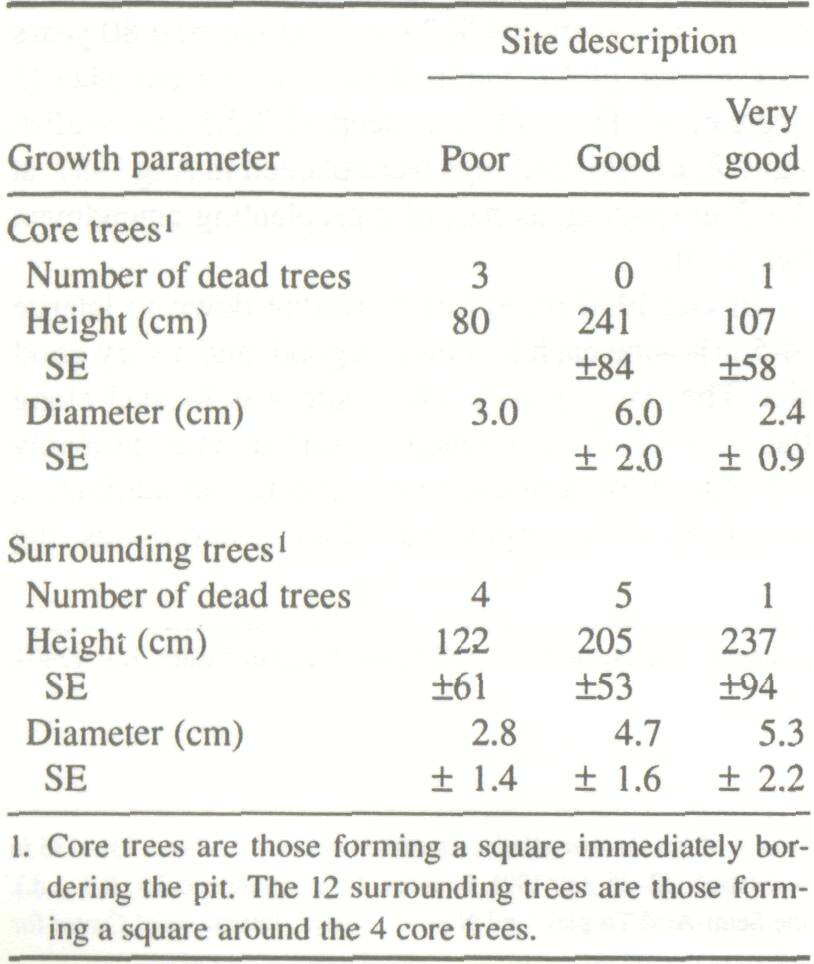
The field trial was located at the ICRISAT Sahelian Center, near Niamey, Niger (13N 2ºE), on a slightly undulating eolian sand plain. Annual rainfall in Niamey has averaged 562 mm over the past 80 years (Sivakumar 1986). the rainy season is from May to September. The soil is a deep (4.2-5.1 m) reddish sand. F. albida seedlings were planted in Aug 1987, at 2x2 m spacing, as part of a preplanting amendment field trial. In Oct 1990 three pits w (4-5 m)one each in a poor, site. The 'very good growth the edge of an abandoned t constructed by a Macroterm trench 30 m in length and from the poor to the good growth area. A second 15-m long trench was dug parallel and 25 m away from the first, leading from the center of the abandoned termite mound to a second poor growth area. In the three pits, full profile descriptions were made (FAO 1977) and all horizons (8-10 per profile) were sampled for physical, chemical, and micro-morphological analysis. Evidence of biological activity was mapped to scale on graph paper, down to a depth of 3.5-4.0 m using a 1 x 1 m metal frame which was divided into 10 x 10 cm squares and hung against the pit face. From these maps, the relative areas of various types of biological activity (roots, termite activity, and activity by other fauna) were estimated. Similar biological activity maps and calculations were made along the full length of the two trenches. Depths to horizon boundaries were determined, and samples were taken at 9 depths every meter (second trench) or every two meters (first trench) for determination of pH (in water), texture and bulk density. Around all pits and trenches tree height or length of the longest branch and diameter at 10 cm were measured.
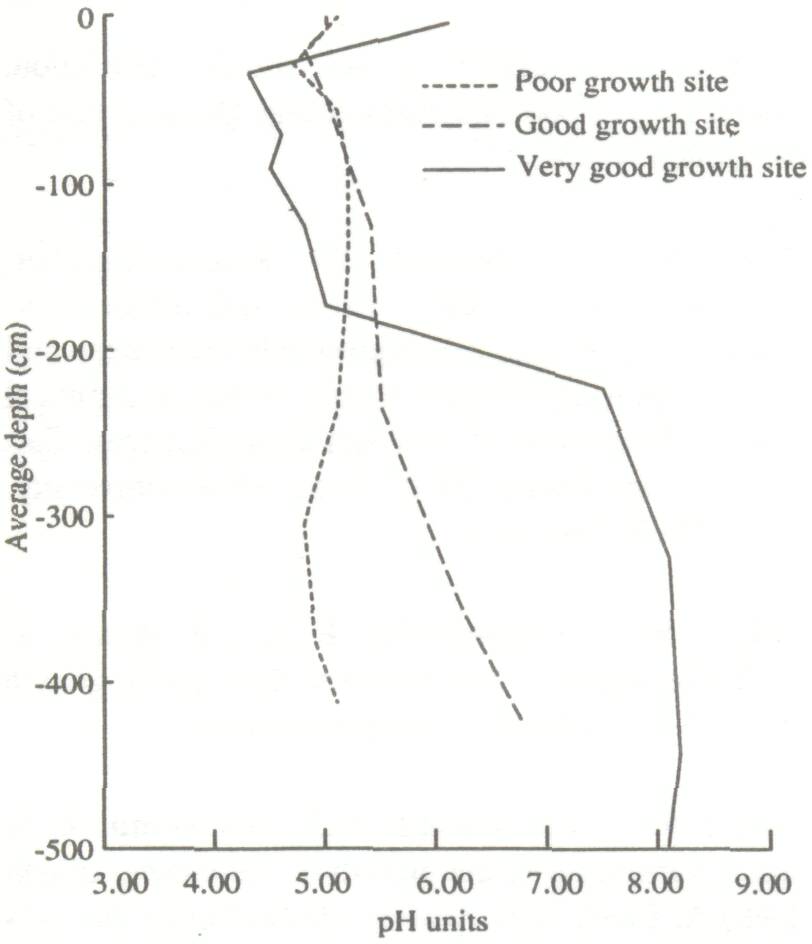
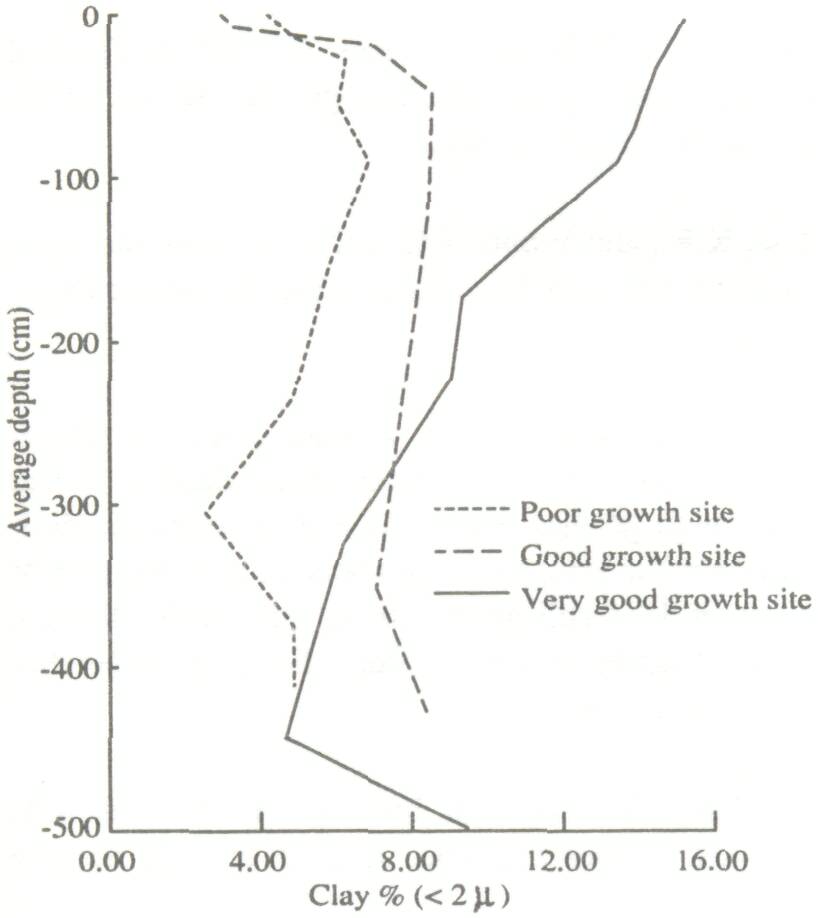
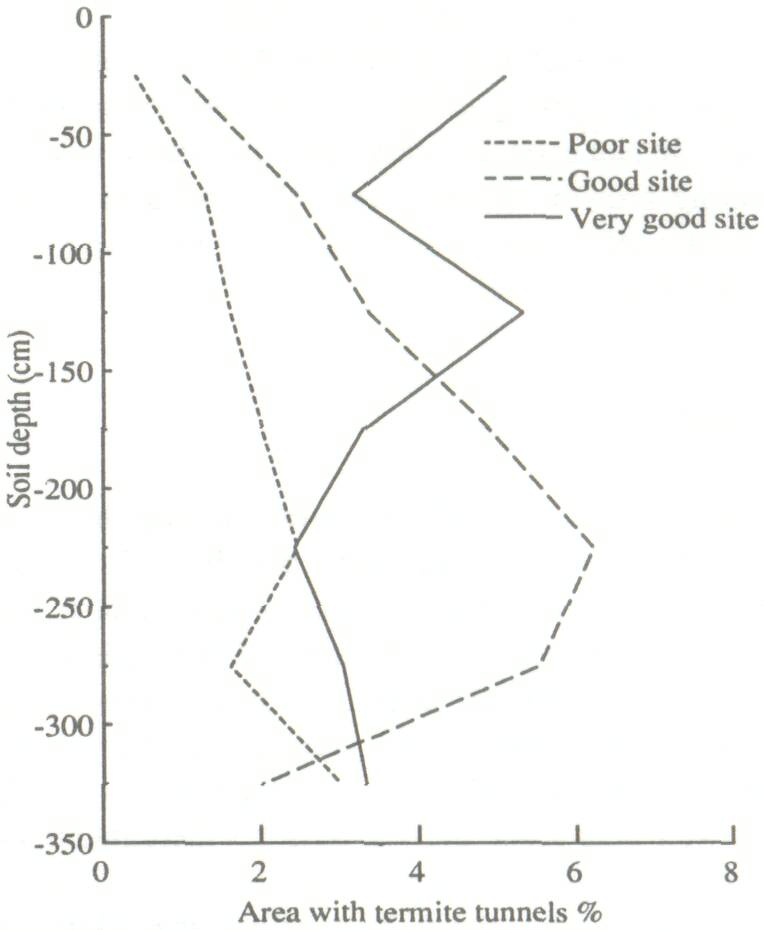
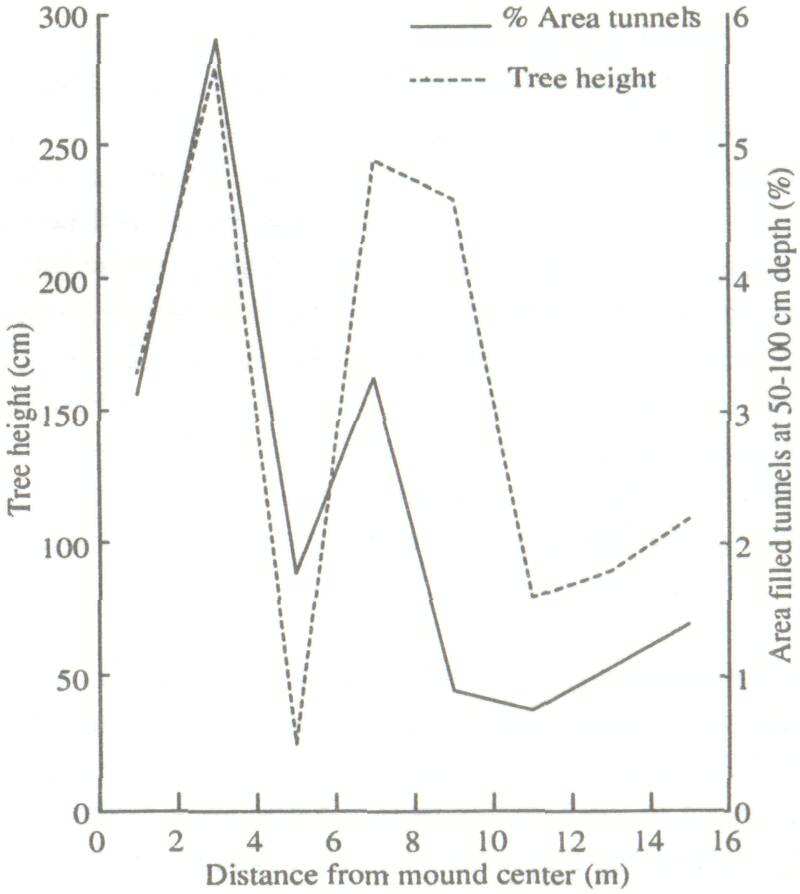
Height and diameter for the four trees immediately surrounding the termite mound are low (Table 1). This is because the growth conditions at such sites are often unfavorable due to high bulk density and low infiltration capacity (Lal 1987). Areas around recently abandoned mounds are often favorable to growth, because of runoff from the degraded mound as well as increased fertility. This is reflected in the growth figures for the 12 trees immediately around the mounds. Except for the surface crust, there was a relatively high organic carbon content to a depth of 3 m under the termite mound. The poor growth area had low organic carbon and the good growth area was intermediate. Bray-1 P showed the same trend, as did total N (weakly) and free Fe. The pH (H2O) also followed this pattern, except for a completely worked zone immediately under the old mound (to a depth of 1.9 m), which was very acid (Fig. 1). Similarly, clay contents are highest under the old mound, and lowest under the poor growth (Fig. 2). All these trends more or less coincide with termite activity (mostly back-filled tunnels), particularly in the top 1.5 m of the soil (Fig. 3). The high level of termite activity at 2 to 3 m depth under the good growth area may have had a major effect on tree growth because the tunnels were old and their beneficial effects could be in decline. In support of this, tunnels higher up in the profile were leached and already disappearing. Alternatively, the tunnels might have been buried under more recent eolian deposits. It is also possible that tunnels under this site were made by a different less beneficial termite species. Trenches At the time of writing only data from the trench leading away from the abandoned termite mound were available. The properties of the top half-meter of soil had no major effect on growth of F. albida. This is not surprising, as the seedlings were put into plant holes 40 cm deep and would have very quickly developed a taproot going straight down. We found good correlations, however, between tree height and the relative presence at 50-100 cm of backfilled termite tunnels (r = 0.66, Fig. 4). There were also strong correlations between percentages of backfilled tunnels at 50-100 cm or 100-150 cm, and pH in water at depths of 50 m (r = 0.60), 70 m (0.86), 90 m (0.88), 110 m (0.73), 130 m (0.75), and 150 m r = 0.81). Termite activity and bulk density were correlated only at 50 and 70 cm depth (r = 0.57-0.82). The positive correlation between bulk density at 50 cm depth and tree growth probably has no functional basis; we can conclude only that bulk density was not limiting growth. But bulk densities found in the old mound itself, mostly over 1.60 g cm-3 in the top meter, probably did limit growth.
These data correspond well with findings by other authors that termites can have a positive influence on soil fertility by increasing soil organic matter and clay contents. It might be argued that termites may have a preference for sites that already have better fertility characteristics, e.g., sites that are less leached and have a higher pH. If this was the case at our field site, we would argue that the termites there had made a relatively good site even better. The data also support the conclusions of Geiger et al. (1992), that preexisting soil fertility affects the growth of F. albida and that the so-called 'albida effect' is, to a considerable degree, due to fertility differences that precede the growth of the tree. It appears that, at least in the situation studied, the two findings can be combined by saying that certain termite species can positively affect the growth of F. albida by improving soil physical and chemical fertility. Perhaps their effects on soil biological fertility and soil organisms (bacteria, mycorrhiza, nematodes) should be studied as well. As termites can have a beneficial effect on soil fertility, their well-being should be kept in mind when evaluating agricultural production systems. Tillage has been shown to reduce termite activity (Pierri 1989; Kooyman and Onck 1987); frequent broad acre soil tillage, by affecting termite activity, may in the long term detrimentally affect soil fertility and thus growth of F. albida as well as profitability of agroforestry systems in which F. albida plays a part. Acknowledgment. The trenches were dug by local laborers under the supervision of Mahamadou Djibeye, who also measured the trees. The chemical analyses of the samples from the pits were performed by staff of the Department of Soil Science and Geology, Agricultural University, Wageningen, managed by T. van Mensvoort. Diafarou Amadou diligently analyzed all the pH samples. Ousmane Youm dis-cussed termite identification with us. To all these people, our sincere thanks.
FAO. 1977. Guidelines for soil profile description. Rome, Italy: Food and Agricultural Organization of the United Nations. Geiger, S.C., and Manu, A. 1991. Relationships between soil surface characteristics and millet (Pennisetum glaucum (L) R.Br.) growth in the plateau and valley region of Western Niger. Tropsoils technical report. Department of Soil and Crop Sciences. College Station, Texas, USA: Texas A&M University. (Limited distribution.) Geiger, S.C., Vandenbeldt, R.J., and Manu, A. 1992. Preexisting soil fertility and the variable growth of Faidherbia albida. These proceedings. Geiger, S.C., Vandenbeldt, R.J., and Manu, A. In press. Variability in the growth of Faidherbia albida (Del.) A. Chev. (syn. Acacia albida (Del.)): the soils connection. Soil Science Society of America Journal. Kooyman, C., and Onck, R.F.M. 1987. The interactions between termite activity, agricultural practices and soil characteristics in Kisii district, Kenya. Thesis, Agricultural University, Wageningen, Netherlands. Lal, R. 1987. Termites. Pages 337-421 in Tropical ecology and physical edaphology (Lal, R., ed.). Chi-chester, UK: John Wiley. Lee, K.E., and Wood, T.G. 1971. Termites and soils. London, UK, and New York, USA: Academic Press. 251 pp. Manu, A., Geiger, S.C., Berrada, A., and Scott-Wendt, J.W. 1990. Microvariabilité dans le Sahel: Un apercu général. Pages 117-128 in Actes du séminaire sur l'aménagement des sols, la conservation de l'eau et la fertilisation, 20-24 Feb 1989, Tahoua, Niger. Niamey, Niger: Institut national de recherches agronomiques du Niger. Miedema, R., and Van Vuuren, W. 1977. Termite mounds in Sierra Leone. Journal of Soil Science 28:112-124. Pieri, C. 1989. Fertilité de terres de savanes. Paris, France: Ministère de la Coopération/Institut de recherches agronomiques tropicales et des cultures vivrières. 444 pp. Scott-Wendt, J.W., Chase, R.G., and Hossner, L.R. 1988. Soil chemical variability in sandy Ustalfs in semi-arid Niger, West Africa. Soil Science 145: 414-419. Sivakumar, M.V.K. 1986. Climate of Niamey. Progress Report no. 1. B.P. 12404, Niamey, Niger: ICRISAT Sahelian Center.
1 Department of Soil Science and Geology, Agricultural University, Wageningen, The Netherlands. (ICRISAT), Niamey, Niger. 2 ICRISAT Sahelian Center, B.P. 12404, Niamey, Niger. ICRISAT Conference Paper no. CP 787. Brouwer, J., Geiger, S.C., and Vandenbeldt, R.J. 1992. Variability in the growth of Faidherbia albida: a term Faidherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey Patancheru, A.P. 502 324, India: International Crops Research Institute for the Semi-Arid Tropics; and Nairobi Research in Agroforestry. |