An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 3 Genetics, Provenance Trials, and Vegetative Propagation Invited Paper The Genetics of Acacia albida (syn. Faidherbia albida) H. Joly1 Abstract
Introduction Acacia albida (syn. Faidherbia albida) is a diploid (2n=26) species (Atchison 1948) although a tetraploid form has been seen in Israel. African acacias are for the most part, however, polyploid. The classing of A. albida in the genus Acacia has been discussed for many years. In 1934, Chevalier suggested placing the species in a monospecific genus Faidherbia. Since then, several works (Vassal 1967 and 1969; Guinet and Lugardon 1976; Brain 1987, Dreyfus and Dom-mergues 1981) have shown that A. albida possesses a combination of unusual characteristics among acacias. Several authors have therefore adopted the name Faidherbia albida. Whether this is the definitive taxonomic position of this species is not yet clear; certain authors consider that the genus Faidherbia is part of the family Acacia whereas others state that it would be more appropriately classed in Ingeae.
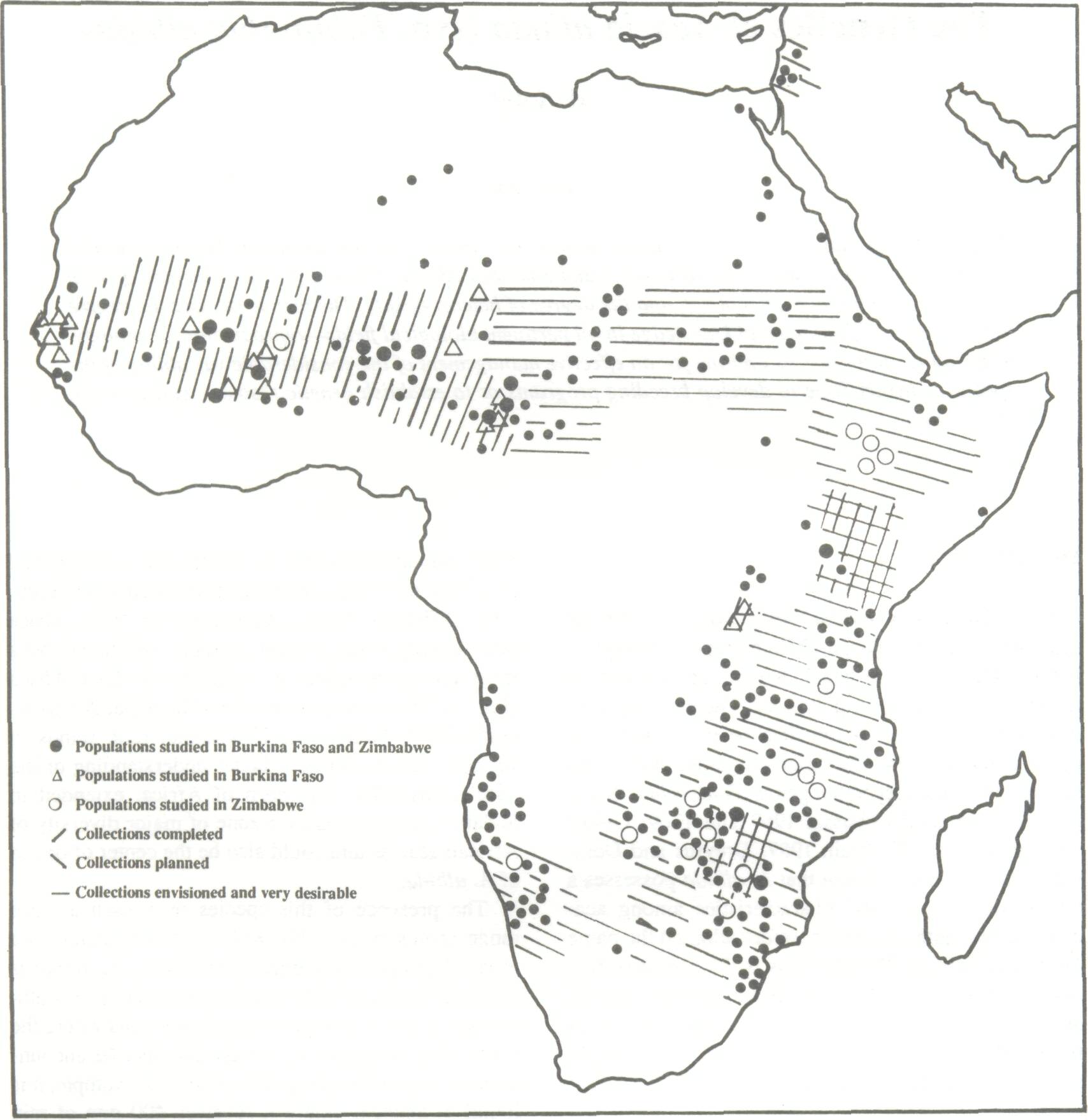
The range of A. albida extends throughout Africa from Senegal to Ethiopia, then southward to Zimbabwe and to the northern part of the Republic of South Africa. It is also found in Namibia and Angola. Relic populations exist in Israel and Lebanon and fossilized trees have been found in the Hoggar Mountains (Wickens 1969). Populations in West Africa have already been, at least partially, collected and a large collection effort is underway in East Africa (Fig. 1). These collections should be expanded to include Chad, Sudan and, if possible, relic stands in various areas to permit a better understanding of the species diversity. The horn of Africa, extended to Sudan, is most probably a zone of major diversity of African acacias and could also be the center of origin of A. albida. The presence of this species over such a large range cannot be possible without its adaptation to a great diversity of ecological conditions. A. albida is generally found on alluvial soils particularly in southern Africa, but it also grows on poorer soils where the water table is deeper. A. albida can also be encountered in very different rainfall zones; for example, it is found in Senegal where it receives 400 mm of rain annually during a 3-month rainy season as well as in Zimbabwe and Burundi where 1000 mm of rain falls in a bimodal pattern. It is also found at altitudes varying from sea level to 2000 m in Ethiopia (CTFT 1988). It was thought until recently that western and eastern African populations were limited to river beds and that only western African populations were associated with agriculture; such stands were, however, observed recently in Malawi (Fagg and Barnes 1990). In the Sudano-Sahelian zone, A. albida is characterized by a defoliation in the rainy season and by a taproot system. This is not systematic throughout its remaining range. Immature trees in southern and eastern Africa may remain green throughout the year and, in certain zones, the trees are completely defoliated, as in the desert of Namibia (Fagg and Barnes 1990). A. albida may not always be taprooted; shallow-rooted trees have been observed in Senegal and throughout certain eastern African stands.
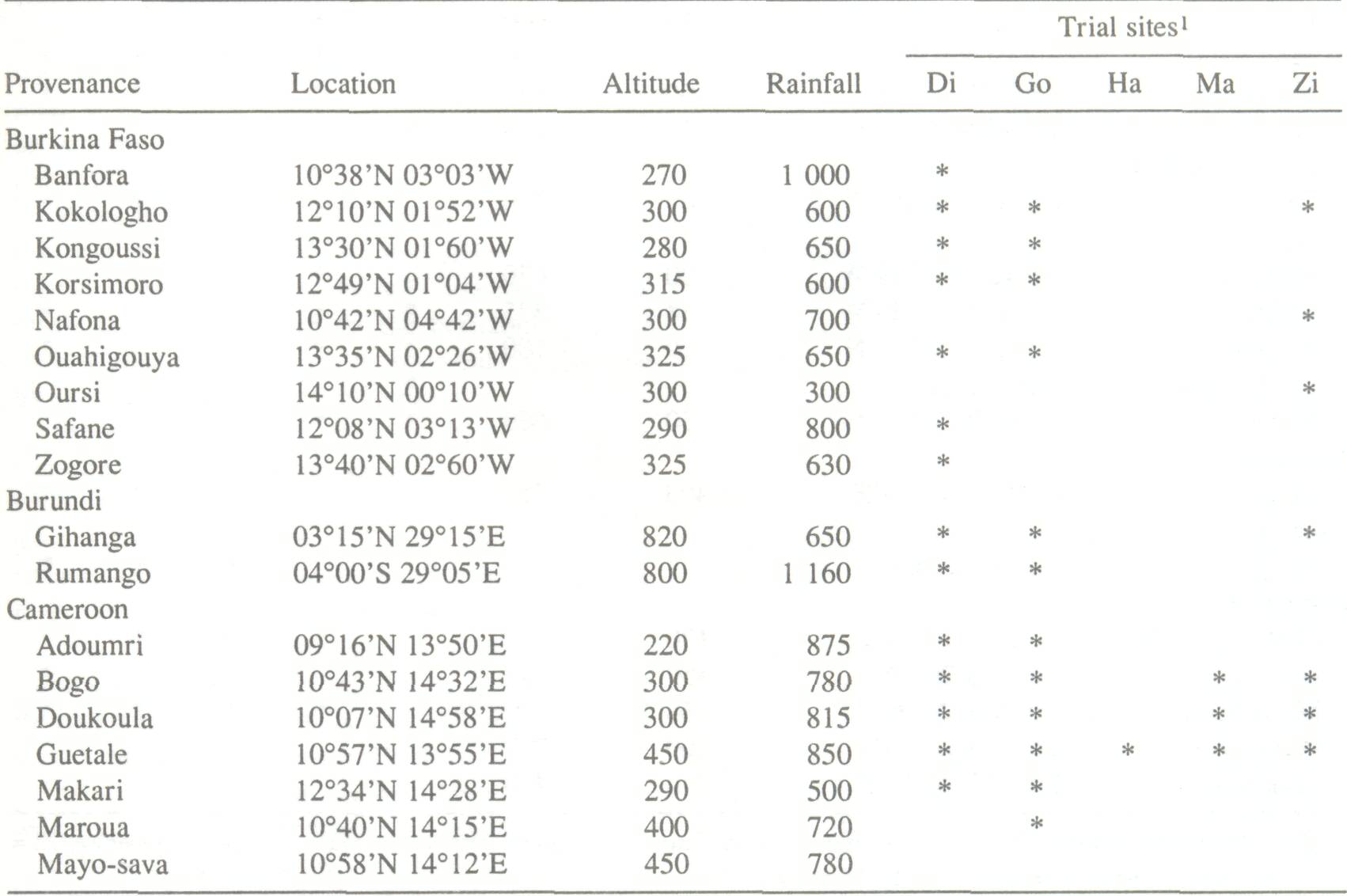
Besides the adaptation of A. albida to such diverse ecological conditions, there is considerable morphological variation of characters such as tree or pod form. In support of development activities, breeding programs and gene resource conservation strategies for A. albida must quantify the variability and determine how much of this is due to genetic or environmental differences. The spatial structure of this genetic diversity must also be studied. Despite the importance of A. albida, very little information exists on the genetic diversity of numerous traits of agronomical or pastoral interest. Since pertinent traits have not yet been defined, there is nothing known regarding the range of their variability and their heritability. Phenotypic Variability Numerous trials have been planted as part of development programs in Senegal since 1964, and in Niger in 1973 and 1974 (Montagne 1984; Cazet 1987). However, the first provenance trials were not installed until 1985 in Burkina Faso and 1986 in Zimbabwe (Table 1). The Burkina Faso trials were located in Dinderesso (950 mm rainfall) and Gonse (800 mm) utilizing seed origins from Burkina Faso, Mali, Niger, Senegal, more central provenances (Cameroon) and eastern Africa (Burundi, Ethiopia, Zimbabwe). Some of these same seedlots were raised in greenhouses in Zimbabwe for other traits. One trial, which focused more on agroforestry was laid out in Cameroon in 1984 to evaluate the tree's impact on rotated crops (sorghum, cotton, groundnuts) (Peltier and Eyog-Matig 1988) and to compare different provenances.
The Burkina Faso trials focus only on survival traits and growth from 6 months to 5 years. The Zimbabwe trials involved younger stock and focused on production traits and fodder quality. In both trials, provenances of eastern and southern Africa showed better initial growth than those of western Africa. Yet, the Burkina Faso trials showed that these provenances were poorly adapted to a Sudano-Sahelian climate because the mortality rate at Gonse approached 100% by the second dry season. In Dinderesso, where the annual rainfall is greater and the rainy season longer, the Burundi provenance remains vigorous (Billand and de Framont 1990), but this provenance also failed in Mouda, northern Cameroon (Peltier and Eyog-Matig 1988). It should be noted that these results were obtained at a juvenile stage of the plants and may be partially due to a difference in seed size. Seeds of southern African provenances are two to three times larger than seed of western African provenances. Sniezko and Stewart (1989) also show that western African provenances, consisting of 3-month old noninoculated stock in a nursery, had poorer nodulation than eastern and southern African provenances. Root:shoot ratios averaged 50% for all the provenances, and were slightly higher for western African provenances. These initial results are interesting for evaluating response of different seed origins under varied ecological conditions; they do not, however, provide much useful information on the heritability of the traits studied and hence on the potentials of selection for these traits in a breeding program. Progeny trials have recently been set up in Burkina Faso to evaluate variation of traits. These trials include three provenancesKagnobon (Senegal), Kongoussi (Burkina Faso), and Matameye (Niger)which are all represented at the other trials in Burkina Faso. The first trial was laid out in 1987 for the Kongoussi provenance. Heritability at 6 months was 38% for height and 22% for diameter (at the root collar); at 18 months, heritability was 19% for height and 16% for diameter (IRBET-CTFT 1989). All these results focus on the differences between western African provenances and those of eastern Africa (Table 2); it is difficult to evaluate which part can be attributed to genetics in these differences. Together, these measurements describe growth traits and form at the juvenile stage. It would be desirable to expand this type of study to include essential physiological differences of the species at maturity, such as shedding of leaves in the rainy season or water utilization.
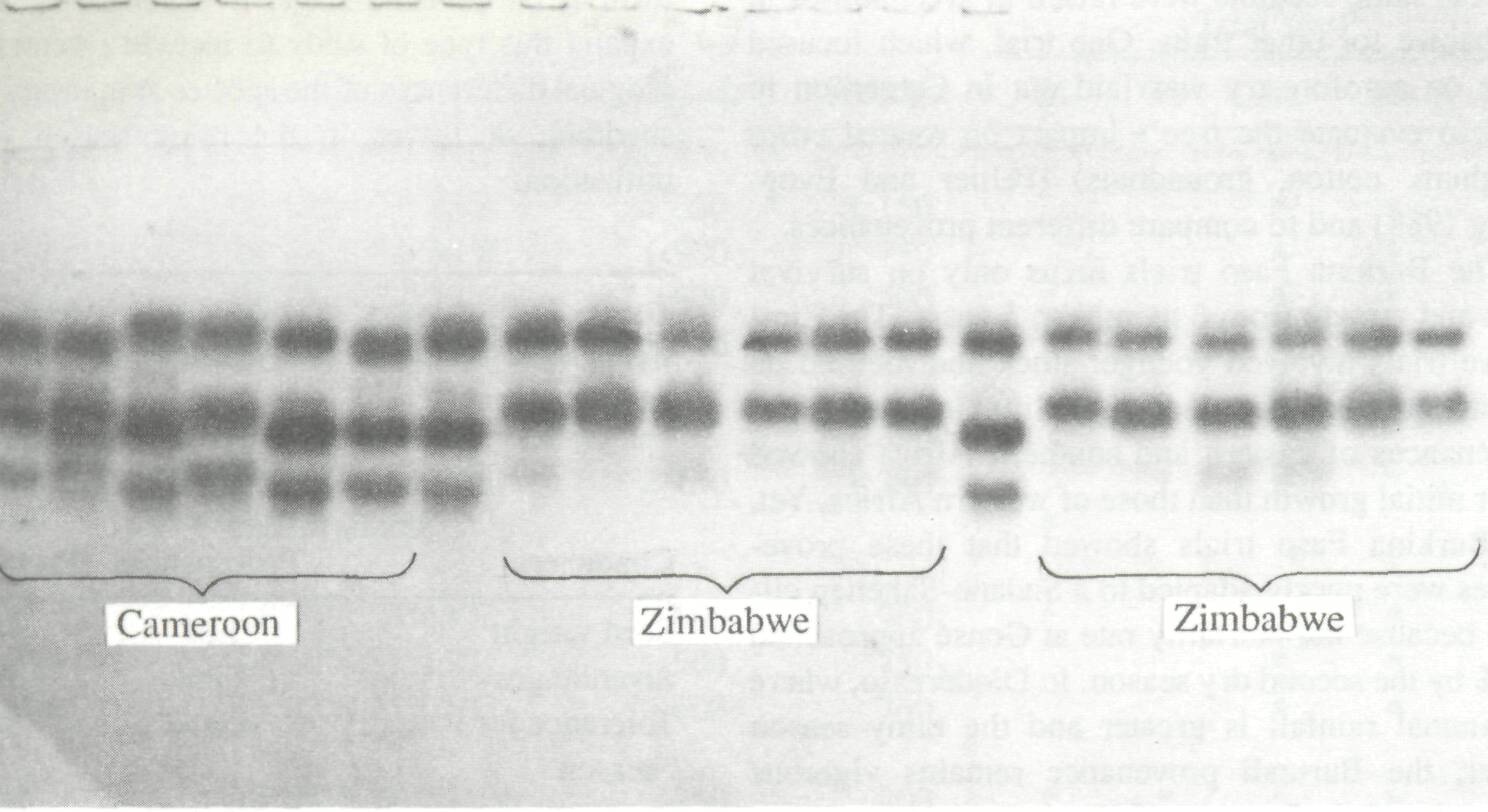
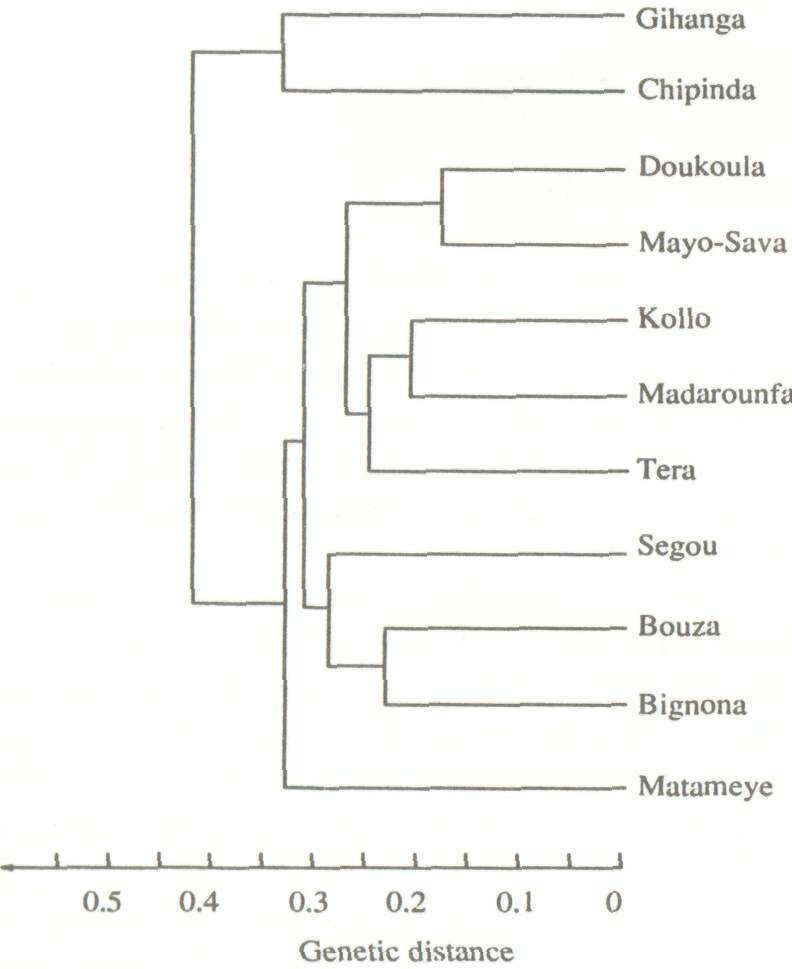
The expression of important agronomic, morphological, or physiological characteristics depends on the interaction of environmental and genetic parameters. Thus, complementary studies of variability of species that emphasize purely genetic traits which are not directly susceptible to environmental pressures, are useful. A study of the enzymatic variability of A. albida is being undertaken in France (Danthu and Prat 1991; Zeh-Nlo et al., in press) and in the UK at the Oxford Forestry Institute. Trials have been conducted with eleven provenances, including three from western Africa (Mali, Niger, and Senegal); two from Cameroon and two from eastern Africa (Burundi and Zimbabwe). Eleven enzymatic systems representing 16 loci were studied using 20 trees per provenance. This study has conclusively demonstrated the great diversity of A. albida, particularly between eastern and western African provenances (Fig. 2). A large separation is evident in eastern African populations relative to others vis-a-vis the absolute genetic distance (Fig. 3) (Joly and Zeh-Nlo, in press). A study of stands in the region surrounding Sudan should determine whether western African populations are derived from the eastern populations or vice versa ora, more likely propositionwhether they are derived from a common source located in this zone.
Reproductive Biology and Organization of Genetic Diversity Sexual Reproduction The reproduction of A. albida is poorly known. Its pollen is a polyad of 30 monads (Guinet 1969). The species is pollinated by entomophilous means. Recent studies (Tybirk 1991) show that the pollinators belong to numerous families of which Scoliidae and Eumonidae (hymenopterae), and Lycanidae and Danaidae (leptidopterae) are the most frequent. Isolated trees produce seed, therefore it appears likely that A. albida does not possess a strict self-incompatibility system. However, the level of self-fertilization is not known. Flowering of individual trees is often not uniform and, therefore, the rate of self-fertilization within a stand may vary from tree to tree through the flowering stage. The study of enzymatic systems may also allow estimation of the rate of a tree's self-fertilization and the level of a population's homozygosity. Studies of enzymatic systems done at ENGREF show that all A. albida populations studied reveal a deficit in heterozygotes one would not expect in a completely panmictic population. It should be noted that eastern African populations present a larger deficit than those of western Africa. Vegetative Reproduction Under certain conditions, A. albida can reproduce by vegetative means. Establishments by runner cuttings were monitored in Israel (Karschon 1976) where, despite considerable flowering, each pod produced contained no more than one or two very small seeds. It is probable that a combination of conditions for good germination did not exist because of cold temperatures during the rains. In certain areas of West Africa the regeneration of natural parks could also be done at least partially by runner cuttings. Studies of the spatial structure of the genotypes obtained by electro-phoresis for stand establishment are planned in Senegal; this will permit evaluation of vegetative multiplication in regenerating tree stands (Personal communication, A. albida Research Program, Bernard Dreyfus, Coordinator, ORSTOM, Dakar). Organization of Genetic Diversity The biology of reproduction and means of seed dispersion defines the spatial nature of genetic diversity. Entomophilous pollinization in A. albida suggests that pollen dispersion is of little consequence. However, the pods are consumed by livestock and wildlife and can therefore be disseminated over several hundreds of square kilometers, often to areas where cattle rest. The practice of selection by the farmer in his field can also affect the level of order of genetic diversity of A. albida. Preliminary electrophoretic studies of this hypothesis show that the differences between populations is a considerable (FST=0.23) part of total variability (FIT=0.44) (Joly and Zeh-Nlo, in press).
Designing a breeding strategy for a multipurpose species is always difficult when traits to be selected are numerous. The simultaneous selection of several traits may prove to be impossible if these traits are negatively correlated. Furthermore, the highly diverse conditions under which the species is grown should be taken into consideration because lines bred for maximum growth on an optimum site may not be sufficiently hardy for adaptation elsewhere. Clearly, the evaluation of genetic diversity is necessary in order to define breeding programs as well as to establish a genetic resource conservation strategy. For that reason basic knowledge on the species must be compiled and several tools well understood in advance. Some of these are described below. Controlled Crossings The mastery of controlled crossings is important for any breeding program. Pod production from such crossings is one of the principal interests in A. albida breeding; a thorough knowledge of flowering and fruit-bearing process is therefore particularly important. Vegetative Propagation Several cutting trials are under way in Burkina Faso (Bonkoungou et al. 1988) and in Senegal (Danthu, personal communication). A few grafting trials have also been undertaken in Burkina: the initial results have not been encouraging. Trials have also been carried out in order to develop micropropagation techniques from juvenile crown material (Duhoux and Davies 1985) and adult stage crown material (Gas-sama and Duhoux 1987). A new method which appears very promising involves multiplication by root fragments. These initial efforts are yielding good results with juvenile material. Work with cuttings is in progress (Duhoux and Gassama, personal communication). It is likely that material will soon be available for clonal testing and for establishing conservation orchards of certain endangered stands.
Studies are under way to evaluate the degree of nodulation under natural conditions and to study rhizobium-mycorrhiza interactions and their effects on A. albida growth. The works of Lajunie et al. (1990) show that A. albida nodulates with Bradyrhizobium rather than with Rhizobium as most African acacias do. In Burkina Faso, a strong interaction has been shown between Rhizohium stock and mycorrhizae (Dianda 1990). A breeding program is being developed in Burkina Faso by IRBET-CTFT with the aim to produce sufficiently homogenous planting material and better initial growth. A seed orchard will be established and clonal testing is planned. Studies will be carried out on the floral biology of A. albida with the focus on the tree-Rhizobium-endomycorrhiza interaction (Bonkoungou et al. 1988). To improve such activities, further studies should be carried out on water use and the phenological rhythm of A. albida to provide better selection criteria.
Pooled results show that despite African farmers' and herders' longstanding interest in this species, and the importance placed on it by the scientific community, there is a lack of basic knowledge on A. albida. The role of A. albida in agroforestry systems, its impact on the water balance, and on mineral element cycling have not been elucidated. Leaf fall during the dry season is an important characteristic. It remains to be confirmed whether the phenomenon is due to environment or genetic factors. This lack of knowledge makes it difficult to choose useful traits for selection. Furthermore, negative or positive correlations may exist between these physiological characteristics as well as other important characteristics such as initial growth, production, and quality of fodder produced. Ignorance of adaptive mechanisms of A. albida also makes in situ protection of genetic resources difficult since the impact of environmental modifications on these mechanisms and on the reproductive systems of A. albida is ignored. The need for effective applied and sustained research cannot be stressed enough. All integral development plans for A. albida are hampered by failure to recognize the biological nature of this species.
Atchison, E. 1948. Studies in the Leguminosae: II. Cytogeography of Acacia (Tourne.) L. American Journal of Botany 35(10):651-655. Billand, A., and de Framont, H. 1990. Variabilité génétique de Faidlierbia albida (Syn. Acacia albida) en essais de provenances au Burkina Faso. Presented at the Symposium on Physiologie des arbres et arbustes en zones arides et semi-arides, 26 Mar to 6 Apr 1990, Nancy, France. Bonkoungou, G.E., Ouedraogo, S., Dianda, M., and Billand, A. 1988. Acacia albida Del. en agroforesterie: stratégie d'amélioration du matériel végétal. Presented at the Journées scientifiques universitaires, Ouagadougou, Burkina Faso, 31 May-4 Jun 1988. Ouagadougou, Burkina Faso: Centre national de la recherche scietifique et technique. 22 pp. Brain, P. 1987. Immunology and phylogeny: a preliminary study of Acacia. South African Journal of Science 83:422-427. Cazet, M. 1987. La régénération artificielle de Faidherbia albida en zone sahélienne: plantation ou semis direct? Premiers résultats de l'expérimentation conduite à Thiénaba (Sénégal) en 1986. Dakar, Senegal: Direction des recherches sur les productions forèstieres. 49 pp. (Limited distribution.) Chevalier, A. 1934. Nouvelles observations sur quelques Acacias de l'Afrique occidentale. Revue de Botanique Appliquee 14:875-884. CTFT (Centre technique forestier tropical). 1988. Faidherbia albida (Del.) A. Chev. (Synonyme Acacia albida Del.) Nogent-sur-Marne, France: CTFT. 72 pp. Danthu, P., and Prat, D. 1991. Study of the genetic variability by means of isoenzymes in Faidherbia albida: preliminary results. Pages 225-226 in Biochemical markers in the population genetics of forest trees. (Fineschi, S., Malvolti, M.E., Cannata, F., and Hattemer, H.H., eds.). SPB Academic Publishing bv. Dianda, M. 1990. Etude de la double symbiose Rhizobium-mycorhizes VA chez Acacia albida au Burkina Faso. Presented at the Symposium on physiologie des arbres et arbustes en zones arides et semi-arides, 26 Mar to 6 Apr 1990, Nancy, France. Dreyfus, B.L., and Dommergues, Y.R. 1981. Nodulation of Acacia species by fast- and slow-growing tropical strains of Rhizobiuni. Applied and Environmental Microbiology 41(l):97-99. Duhoux, E., and Davies, D.W. 1985. Caulogenese a partir des bourgeons cotyledonaires d' Acacia albida et influence du saccharose sur la rhizogenese. Journal of Plant Physiology 121:175-180.
Fagg, C.W., and Barnes, R.D. 1990. African acacias: study and acquisition of the genetic resources. Final Report on Overseas Development Administration Research Scheme R.4348. UK: Oxford Forestry Institute. 170 pp. Gassama, Y.K., and Duhoux, E. 1987. Micro-propagation de 1'Acacia albida Del. (Leguminosae) adulte. Bulletin de l'IFAN, T. 46, series A, 3-4:314-320. Guinet, P. 1969. Les Mimosacees. Etude de pa-lynologie fondamentale, correlations, evolution. Insti-tut fran9ais de Pondichery. Travaux de la section scientifique et technique. 293 pp. Guinet, P., and Lugardon, B. 1976. Diversit6 des structures de 1'exine dans le genre Acacia (Mimo-saceae). Pollen spores 18(4):483-511. IRBET/CTFT. 1989. Rapport annuel d'activites 1988. Ouagadougou, Burkina Faso: Institut de re-cherche en biologie et ecologic tropicale/Nogent-sur-Marne, France: Centre technique forestier tropical. (Limited distribution.) Joly, H.I., and Zeh-Nlo, M. (In press). Population genetics of an African acacia: Acacia albida (syn. Faidherbia albida) 2. Structure of the genetic diversity of eleven populations mostly from West Africa. Karschon, R. 1976. Clonal growth patterns of Acacia albida Del. Bulletin of the International group for the study of Mimosoideae 4:28-30. Lajunie, de P., Neyra, M., Dupuy, N., Alazard, D., Gillis, M., and Drefus, B.L. 1990. Diversite des rhi-zobium et specificite de nodulation chez les acacias saheliens. Presented at the Symposium on physiologie des arbres et arbustes en zones arides et semi-arides, 26 Mar to 6 Apr 1990, Nancy, France. Montagne, P. 1984. 'Faidherbia albida': son devel-oppement au Niger. Aix-en-Provence, France: Bois de Feu. 90 pp. Peltier, R., and Eyog-Matig, O. 1988. Les essais d'agroforesterie au Nord-Cameroun. Bois et Forets des Tropiques 217(3)3-31. Sniezko, R.A., and Stewart, H.T.L. 1989. Range-wide provenance variation in growth and nutrition of Acacia albida seedlings propagated in Zimbabwe. Forest Ecology and Management 27:179-197. Tybirk, K. 1991. Report for the field work in Senegal December 1990/January 1991. Denmark: Botanical Institute, University of Aarhus. 4 pp. Vassal, J. 1967. La plantule d'Acacia albida Del. (Faidherbia albida (Del.) A. Chev.). Bulletin de la Societe d'Histoire naturelle de Toulouse 103(3-4):583-589. Vassal, J. 1969. Contribution a l'etude de la mor-phologie des plantules d'Acacia. Acacias africains. Bulletin de la Societe d'Histoire naturelle de Toulouse 105:55-111. Wickens, G.E. 1969. A study of Acacia albida Del. (Mimosoideae). Kew Bulletin 23(2):181-202. Zeh-Nlo, M., Danthu, P., and Joly, H.I. (In press.) Population genetics of an African acacia: Acacia albida (syn. Faidherbia albida) 1. Genetic control of 14 enzymatic systems.
footnote__________ 1. Ecole nalionale du genie rural des eaux et des forets (ENGREF), 14 rue Girardet, FS 4042, Nancy, France. Joly, H. 1992. The genetics of Acacia albida (syn. Faidherbia albida). Pages 53-61 in Faidherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey, Niger (Vandenbeldt, R.J., ed.). Patancheru, A.P. 502 324, India: International Crops Research Institute for the Semi-Arid Tropics: and Nairobi, Kenya: International Centre for Research in Agroforestry. |