An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 2 Uses Invited Paper The Fodder Role of Acacia albida Del.: Extent of Knowledge and Prospects for Future Research M.I Cissé1 and A.R. Koné2 Abstract
Introduction In the semi-arid régions of West Africa, trees play an essential role in the life of rural populations and in traditional agrarian systems. In Sudanian and Sahelo-Sudanian zones, Acacia albida is regularly associated with the most intensively cultivated lands ("Soforo" in the Bamanan dialect; "Lara" in Dogon). According to Pélissier (1979), A. albida stands are critical to the lives of farmers dependent on rainfed cereal agriculture and breeding of domestic animals. A. albida is at the foundation of this agropastoral system. Seignebos (1978) indicates the importance of this species: "Thanks to its reverse [phenology], [A. albida provides] a...microclimate favorable to crops. By its litter, it enriches the soil and permits increased millet and sorghum yields [without requiring] fallow periods. The nutritive value of its fodder is as important as its [fodder role in the dry season]. [And despite intensive exploitation], it tolerates delimbing." In other respects, the seeds, gum, bark, and wood are utilized for many purposesfood, traditional medicine, construction, furniture, canoes, and other domestic usesmaking A. albida the "miracle" tree of the Sahel. This paper summarizes the knowledge compiled on the fodder role of A. albida while focusing on the close relationships that exist between this species and domestic animals. Because of its highly nutritional leaves and fruit, A. albida provides domestic animals with excellent fodder in the dry season; in return, cattle assure its regeneration by pretreating the seeds in their digestive tract and then acting as disseminators.
In West Africa, references to A. albida as a fodder tree are numerous (Curasson 1953 and 1958; Gillet 1960; Adam 1966; Audru 1966; Boudet 1970 and 1972; Diallo 1968; Mosnier 1961 and 1967; Peyre de Fabrègues 1963; Naegelé 1971; Touzeau 1973; Giffard 1974; Le Houérou 1980b). All of these authors agree that A. albida leaves and pods serve as fodder for cattle, sheep, goats, and camels; however, they make no mention of the production of this species. Interest in evaluating fodder production of tree species is recent (Bille 1977 and Poupon 1980 in Sénégal; Nebout and Toutain 1978 in Burkina Faso; and Cissé 1980 in Mali). Such evaluation involves problems linked to:
Besides the pods, which have obvious fodder value, a number of other tree components may warrant attention. The fodder value of tree bark, which is generally of little or no interest to domestic animals, has rarely been evaluated, and never for A. albida. The branches, usually evaluated with the leaves, may have distinct fodder potential and therefore deserve a separate evaluation. The flowers of A. albida do not appear to be much sought after by domestic animals; however, the fruit has fodder potential. The production of A. albida fruit was evaluated in Niger (Lemaitre 1954), Sénégal (Jung 1967) and Sudan (Wickens 1969). Tree leaves, generally the most browsed part of the tree, are an important fodder source. Cissé (1980) evaluated the foliar production of A. albida in Mali, and many other similar studies have been done on other species. Due to the relative abundance of research done on the fodder potential of A. albida leaves and pods, the following discussion will be limited to these two components.
A. albida bears fruit between January and April, a critical period in terms of fodder availability. Fallen fruit are consumed and, as this period progresses, they are knocked down and then collected, and fed to animals or sold in markets. Pods of A. albida mature slowly. In Niger, Bau-mer (1983) found that, under very exceptional conditions, pod development begins between the 2nd and 4th years but in general, fruit-bearing commences around the 15th year. In Sénégal, Nongonierma (cited by Felker 1978) observed that the first pods appear during the eighth year; in Niger, Lemaitre (1954) observed that they appear between the 8th and the 12th year. Reports of A. albida pod production are sometimes unreliable. Based on an average established on twenty 30-year old trees, Lemaître (1954) estimated a production of 6 to 8 kg tree-1 around Zinder in Niger. In Bambey, Sénégal, Jung (1967) estimated fruit production of one tree (crown surface area = 230 m2) at 125 kg. Wickens (1969) harvested 135 kg from a mature tree in Sudan. Evaluating these diverse results, Felker (1978) concluded that "because of marked differences between Lemaître's results and those of others, the absence of information on the method used by Wickens and the serious limitations of Jung's method, we should consider the average pod production per tree and per hectare as unknown." Thus, these data do not have indicative value. The evaluation of pod production, therefore, requires the adoption of a viable method. One method may be to determine whether correlations can be established between pod production and physical parameters, for example, crown size (Lemaitre 1954). Referring to evaluations done on Acacia tortilis in Kenya, Ciss6 (1983), noted positive correlations between pod production and height (r = 0.72), trunk circumference (r = 0.92) and crown area (r = 0.85). Applying these allometric relationships to inventory data would allow evaluation of stand production. To establish allometric relationships, individual tree production was evaluated in the following order: aerial harvesting of pods; estimation of areas over which the pods were gathered; evaluation of collected pod density by counting over an area of 12 x 0.25 m; estimation of pods remaining on the tree in relation to those collected; and evaluation of mass of pods sampled relative to the total number of pods. According to Bille (1977) and Poupon (1980), fruit production fluctuates from year to year among Sahelian tree species. To allow for these variations, the evaluation of pod production should take into account only trees bearing fruit at the time of sampling.
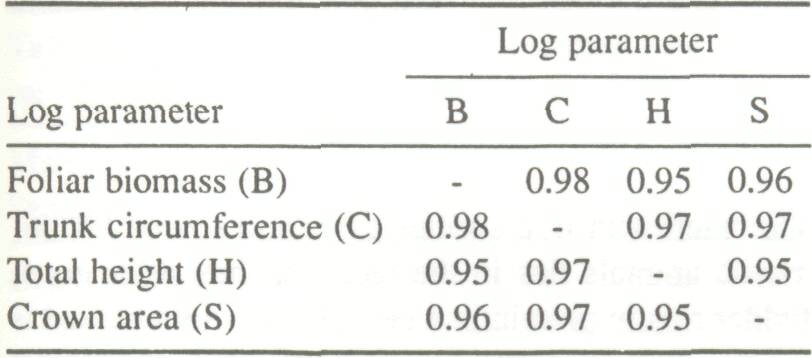
Seasonal Distribution Cissé (1980) compared foliar biomass at peak production (B) using data based on five trees per trunk-circumference class (10-cm classes) with the following parameters:
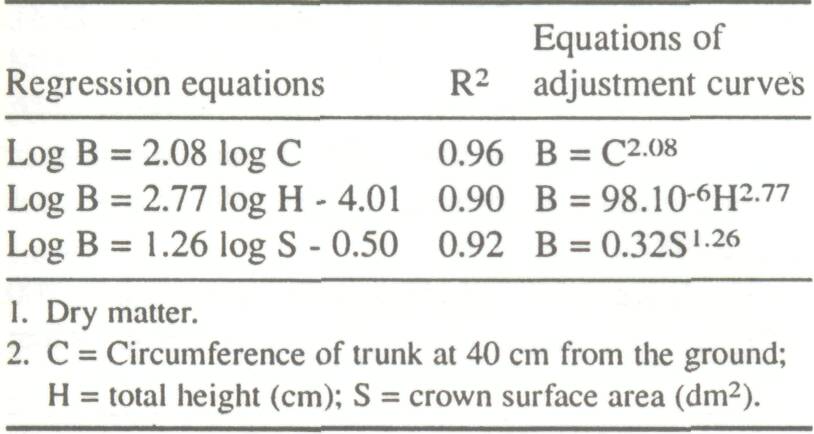
The most significant physical parameter linked to foliar biomass was analyzed using a log-log linear model on data derived from 50 subjects. The correlation matrices (Table 1) indicate strong positive correlations between the physical parameters as well as between each of the physical parameters and foliar biomass. The linear regression is acceptable (0.90 eR2 <0.96), and the resulting adjustment curves indicate an exponential relationship (Table 2).
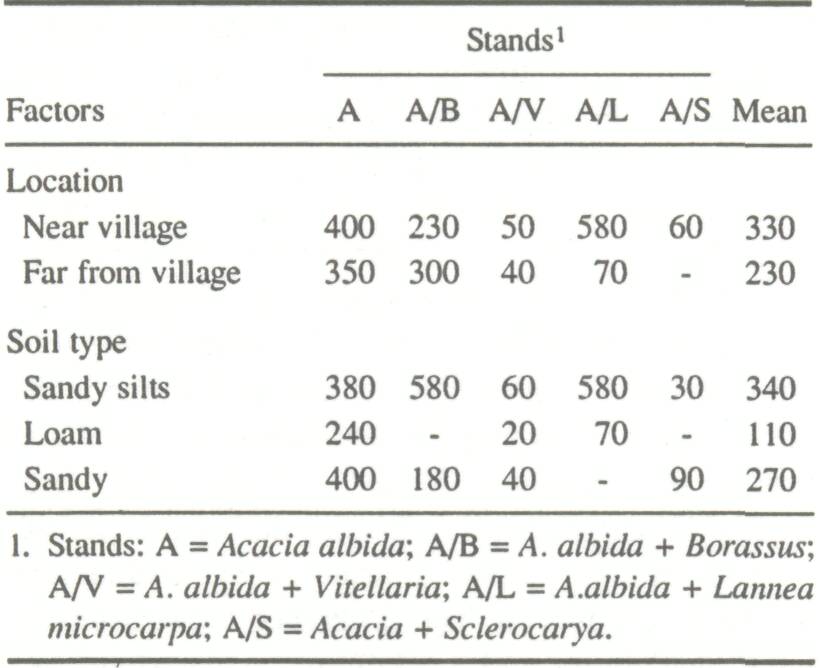
In contrast to other deciduous species, the maximum foliation of A. albida occurs in January during the cold period of the dry season. In the rainy season (July-August), the tree is totally defoliated. Botanists become lost in conjecture and speculation when considering the "aberrant" phenology of this species (Portères 1957; Lebrun 1968; Trochain 1969). However, A. albida owes its importance to this very phenology and provides green fodder during periods of fodder scarcity. Fodder exploitation extends the foliating period of A. albida and enhances foliation in the rainy season. This may be detrimental to crops and provide little benefit to domestic animals, which prefer highly nutritious green grasses during this period. The foliation curves indicate foliage availability on the stump as well. The difference between maximum availability and availability on the stump reflects the extent of defoliation. Foliar Exploitation in Mali Foliar production figures represent foliar biomass of a tree as a function of its physical characteristics. They are utilized in descriptive forest inventories to estimate foliar biomass of a stand. In Mali, A. albida is found in pure stands or in stands associated with Vitellaria paradoxa, Borassus aethiopium, Sclerocarya birrea, and Lannea microcarpa. These occur in the natural stands of Goudo-Mondoro, the Bandiagara-Houbou plateau, in the agroecological zones of the Bas Plateau Bobo, the Koutiala Falo plateau in Falo région, and in the dry Central Delta zone of the Niger River. Forty samples were taken in chosen stands; locations (distances from villages) and soil types were noted. The density, cover, and foliar biomass of the A. albida population in each of these stands were estimated. For every stand, the biomass of A. albida was calculated by summing the contribution of all the clumps from one of the allometric relationships. On the average, A. albida represented 48% of stems and 64% of total cover of these stands and annually produced 300 kg ha-1 of foliar dry matter. These averages, however, mask important differences between stands. As hypothesized, the location of the stand and the type of soil on which it developed also had an impact on foliar biomass production (Table 3).
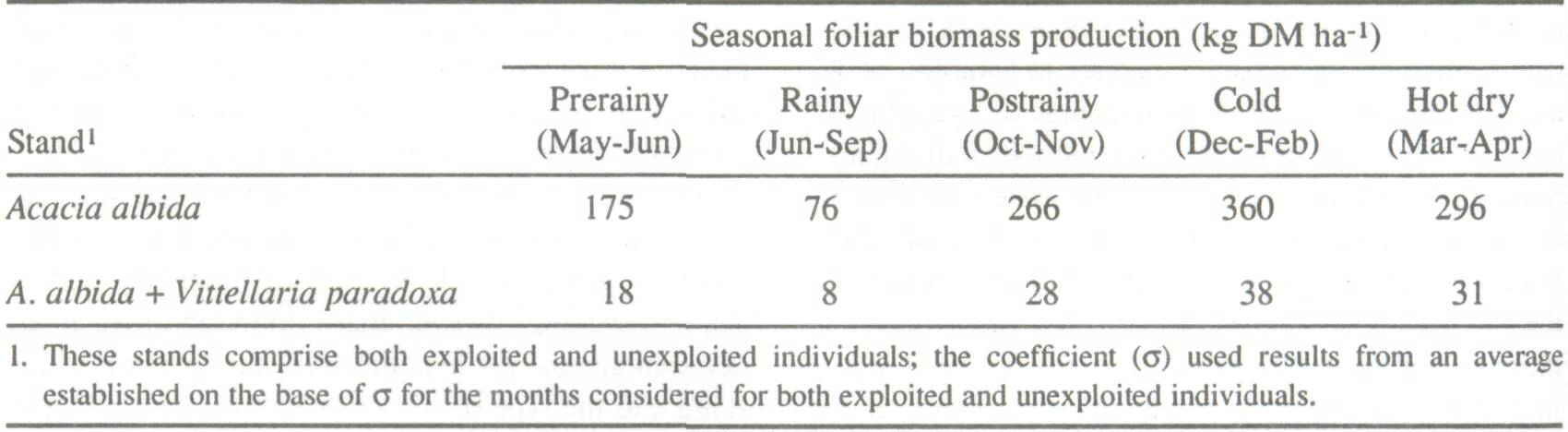
Using maximal biomass estimates, which were derived using allometric relationships, the monthly foliar availability was evaluated by multiplying by the factor σ(σ = bt/bo). As an example, Table 4 shows the seasonal changes and extremes in foliar availability of A. albida by stand and (A. albida + Vittellaria) A/V, which demonstrates the extreme foliar biomass. Effect of Lopping on Production of A. albida Only a small part of foliar production is directly accessible to animals and herders who delimb A. albida. In the short run, this affects fodder production. The frequency and intensity of lopping has a marked im-pact on the foliar production of A. albida (Cissé 1984). For unlopped individuals, repeated pruning during periods of peak average biomass stimulates foliar production. Resulting regrowth is especially vigorous in the first year, but it decreases as exploitation continues. Pruning in January produces more leaves than pruning done in June; however, pruning in June extends the foliation season. A. albida tolerates two prunings per year, but the trees show signs of stress at the end of 6 years. Delimbing was shown to decrease pod production. This is critical not only because of the decrease in fodder potential but also because the pods constitute a commercial commodity for farmers and herders. Rational management of fodder production of A. albida must take into account both pod and leaf production.
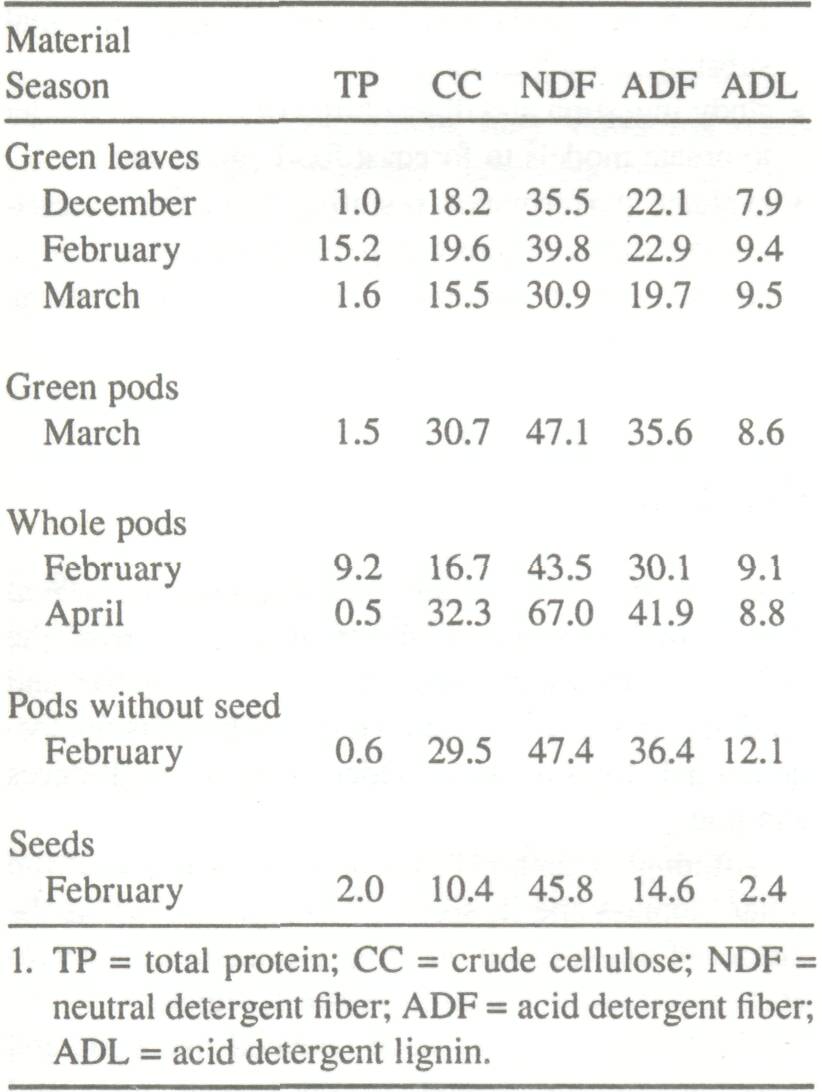
The interest in tree species as a food source for domestic animals lies in the fact that they offer green fodder rich in protein in a period when animal feed is scarce. Behavior studies on cattle and sheep/goat herds in millet agropastoral systems around Niono in southern Sahelian zones of Mali (Dicko 1981) showed that all three types of animal consume browse: about 87% of the goat diet, 36% of the sheep diet, and 11% of the cattle diet are browsed material. The fodder trees are distinguished from forage grasses by their high total protein (TP) content, which ranges from 60 to 250 g kg-1 of dry matter (DM). Some samples have contained up to 330 g kg-1of DM in certain young sprouts and leaves (Rivière 1977; Diagayeté 1981; Koné 1987). Tree fodders are high in lignin content, comprising up to 20% of the mass (Koné 1987). The leaves of A. albida contain an average of 200 g TP kg-1 dry matter (Fall 1978; Le Houérou 1980a; Diagayeté 1981; Koné 1987), the pods contain an average of 150 g, and the seeds an average of 240 to 280 g (A.R. Koné, CRZ personal communication; Wickens 1969, cited by Felker 1978). Following ingestion of A. albida pods, however, only 34% of seeds (which contain a large percentage of TP) are digested (Wickens 1969, cited by Felker 1978). The seed:pod ratio, ranging between 1:2 and 1:8, explains the great variability observed in the chemical composition of these types of fodder (Table 5). The TP includes diverse protein and nonprotein matter which is not discernible in a simple portion of TP. The protein content that is apparently nondigestible can be very high.
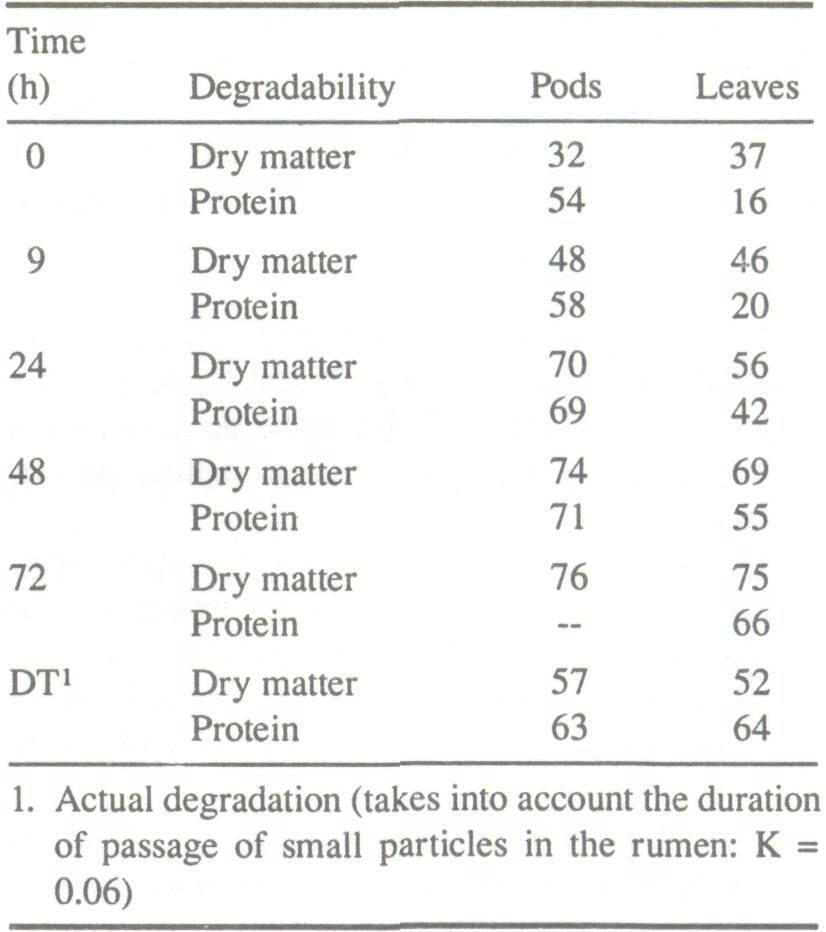
It is difficult to determine the feed value of A. albida just by determining dry-matter, protein, and crude cellulose contents. Estimate of crude cellulose and TP content allows us to evaluate fairly accurately the digestibility of forage grasses with less lignin. Tree species require more detailed analyses of parietal content (method of Van Soest (1983)) and protein content (Koné 1987). Preliminary studies have shown a good correlation between the degradability of TP and the nondegradable protein contents in the acid detergent fiber (ADF) residue (Koné 1987). Measuring the rate of enzymatic breakdown in sacco (in the rumen) of the organic matter and/or proteins is another way of determining digestibility. The results can be compared with those obtained in vitro. The proteins are degraded in the rumen into more soluble products. The extent of degradability varies greatly with the nature of the food and the type of animal. Fall (1978) found that in sheep, solubilization and degradation rate of A. albida pod proteins were higher than those of leaves (Table 6). In goats, however, the intraruminal breakdown of leaf proteins was higher than that of pods (Koné 1987). The dry matter digestibility of A. albida leaves ranges from 14 to 53%. Total protein digestibility varies from 67 to 72%. Dry matter digestibility of whole pods ranges from 50 to 57%; total protein digestibility can reach 73% (Dicko 1979; Fall 1978; Diagayate 1981).
Fodder Efficacy Despite the presence of tannins, A. albidanotably the podsmay be used as a supplement in animal diets because it is rich in protein and digestible energy (77%). Such an energy value would explain the results obtained in Mali (Dicko 1979), where incorporating pods into a sheep feed of sorghum leaves gave an increase in the quantity of dry matter ingested from 40.4 to 61.6 g kg-0.75 in sheep weighing an average of 37.5 kg. The ingestion of leaves by goats was only 41.3 g kg-0.75 per day. In Sénégal, Fall (1978) tested the quality of pods and leaves of A. albida on sheep (weighing on average 22 kg) which received as much rice straw as desired, 100 g of bean cake day-1 per head and bone meal. In addition, one lot of animals received 100 g of A. albida leaves, another 100 g pods, and a third 200 g pods per head. After 92 days, the results were as follows:
This suggests that incorporating pods of A. albida into a low-quality feed enhances digestion without reducing the digestibility of the ration.
Much remains to be learned about the fodder role of A. albida. Because of the multiple-use potential of A. albida, research on its fodder production must take the form of a multi-disciplinary program to improve the contribution of this species to agroforestry systems. In the A. albida-soil-animal association, the mutual benefits can be generally illustrated; however, the returnswhether they be in the form of nutrition, fertilization, or growthhave yet to be fully understood, let alone fully evaluated. To determine the role of fodder in this association, future research should:
Conclusion A species more Sudanian than Sahelian, A. albida derives its importance in fodder production from the fact that it drops its leaves in the rainy season and refoliates in the dry season. Hence, it benefits domestic animals by providing fodder in the form of leaves and pods. Although a method has been developed to evaluate foliar biomass and its seasonal changes as well as the impact of exploitation on foliar production, fruit production has not received the same attention. Feed-value analyses have revealed that pods and leaves are rich in parietal content. Leaves, pods, and seeds contain respectively 200, 150, and 260 g TP kg-1 of dry matter. Antinutritional substances such as tannins limit the digestibility of the abundant protein; however, in spite of the presence of tannins, A. albida componentsparticularly podscan be incorporated into low-quality fodder to enhance ingestion without reducing digestibility. Future research on the role of A. albida as a fodder needs to be done as part of a multidisciplinary research effort that evaluates the contribution of this species to agroforestry systems.
Adam, J.G. 1966. Les pâturages naturels et post culturaux du Sénégal. Bulletin IFAN 28:450-457. Audru, J. 1966. Etude des pâturages naturels et des problèmes pastoraux dans le delta du Sénégal. Etude agrostologique no 15. Maisons-Alfort, France: Institut d'élevage et de médecine vétérinaire des pays tropicaux. Barry, T.N., and Marley, T.R. 1986. Inter-relationships between the concentration of total condensed tannin, free condensed tannin and lignin in lotus sp. and possible consequences on ruminants nutrition. Journal of the Science of Food and Agriculture 37:248-254. Baumer, M. 1983. Notes on trees and shrubs in arid and semi-arid régions. Rome, Italy: Food and Agricultural Organization of the United Nations. 270 pp. Rome, Italy. Bille, J.C. 1977. Etude de la production primaire nette d'un écosystème sahélien. Travaux et Documents no. 65. Paris, France: Institut français de recherche scientifique pour le développement en coopération. Boudet, G. 1970. Pâturages naturels de Haute et Moyenne Casamance (Rép. du Sénégal). Etude agrostologique no. 27. Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Boudet, G. 1972. Etude agrostologique de la cinquième région du Mali (Région de Mopti). Etude agrostologique no. 37. Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Cissé, M.I. 1980. Production fourragère de quelques arbres sahéliens: relation entre biomasse foliaire maximale et divers paramètres physiques. Pages 203-208 in Les fourrages ligneux en Afrique. Etat des connaissances: (Le Houérou, H.N., ed.). Addis Ababa, Ethiopia: International Livestock Centre for Africa. Cissé, M.I. 1982. Variations saisonnières de biomasse foliaire chez quelques ligneux fourragers sahéliens. Program Document AZ 72, Bamako, Mali. Addis Ababa, Ethiopia: International Livestock Centre for Africa. Cissé, M.I. 1983. Disponibilité en gousses d' Acacia tortilis pour la nutrition des petits ruminants dans les ranches Masaϊ de M'birikani et Kimana (Kenya). Program Document AZ 78, Bamako, Mali. Addis Ababa, Ethiopia: International Livestock Centre for Africa. Cissé, M.I. 1984. Synthèse des essais d'ébranchages pratiqués sur quelques ligneux fourragers de 1978 à 1983. Program Document AZ 103, Bamako, Mali. Addis Baba, Ethiopia: International Livestock Centre for Africa. Curasson, M.G. 1953. Arbres, arbustes, buissons et fourrages spontanés divers en régions tropicale et subtropicale. Revue d'élevage et de médecine vétérinaire des pays tropicaux 6(2):37-56, 91-112. Curasson, M.G. 1958. Pâturages et aliments du bétail en régions tropicale et subtropicale. Paris, France: Vigot Frères. Diagayaté, M. 1981. Untersuchungen zurerweiterung der Kenntnisse über den futtwert Westafrikanischer futterpflanzer. Dissertation zur erlangung des grades eines doktors der agrarwissenschaften, University of Hohenheim, Germany. Diallo, A.K. 1968. Pâturages naturels du 'Ferlo sud' (Rép. du Sénégal). Etude agrostologique no. 23. Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Dicko, M.S. 1979. Rapport d'activités 1978-1979 de la nutrition du CIPEA/Mali. Program Document AZ 23, Bamako, Mali. Addis Ababa, Ethiopia: International Livestock Centre for Africa. Dicko, M.S. 1981. La nutrition du bétail du système agropastoral du mil et du riz au Mali. Program Document AZ 64, Bamako, Mali. Addis Ababa, Ethiopia: International Livestock Centre for Africa. Fall, S.T. 1978. Utilisation d'Acacia albida et de Calotropis procera pour améliorer les rations des petits ruminants au Sénégal. Laboratoire élevage et recherche vétérinaire report. Sénégal: Institut Sénégalais de recherches agricoles. 11 pp. Felker, P. 1978. State of the art: Acacia albida as a complementary permanent intercrop with annual crops. Grant no. AID/afr-C-1361. Washington, D.C., United States Agency for International Development. Ph.D. thesis, University of California, Riverside, Cal-fornia, USA. 133 pp. Giffard, P.L. 1974. L'arbre dans le paysage Sénégalais. Sylviculture en zone tropicale sèche. Nogent-sur-Marne, France: Centre technique forestier tropical. 431 pp. Gillet, H. 1960. Etude des pâturages du Ranch de l'Ouadi Rime (Tchad). Journal d'Agriculture Tropicale et de Botanique Appliquee 7(1):1-158. Jung, G. 1967. Influence de l'Acacia albida sur la biologie des sols Dior. Dakar, Sénégal, ORSTOM report. Paris, France: Institut francais de recherche sci-entifque pour le développement en coopération. (Limited distribution.) Koné, A.R. 1987. Valeur nutritive des ligneux fourragers des zones sahélienne et soudanienne d'Afrique occidentale: recherche d'une méthode simple d'estimation de la digestibilité et de la valeur azotée. Ph.D. thesis, University Pierre et Marie Curie, Paris, France. Le Houérou, H.N. 1980a. Le rôle des arbres et des arbustes dans les pâturages sahéliens. Pages 19-31 in Le rôle des arbres au Sahel: Compte rendu du colloque, 5-10 Nov 1979, Dakar, Sénégal. Canada: International Development Research Centre. Le Houérou, H.N. (ed.) 1980b. Les fourrages ligneux en Afrique. Etat des connaissances. Presented at the Colloque sur les fourrages ligneux en Afrique, 8-12 Apr 1980, Addis Ababa, Ethiopia. Addis Ababa, Ethiopia: International Livestock Centre for Africa. Lebrun, J. 1968. A propos du rythme végétatif de 1'Acacia albida. Collectanea Botanica 7(2):625-636. Lemaître, C. 1954. Le Faidherbia albida. Thesis, Niger. 62 pp. Mason, V.C. 1969. Some observations on the distribution and origin of nitrogen in sheep faeces. Journal of Agricultural Science 73:99-111. Milic, B. 1972. Lucerne tannins I: contents and composition during growth. Journal of the Science of Food and Agriculture 23:1151-1156. Mosnier, M. 1961. Pâturages naturels sahéliens: Régions de Kaedi (Mauritanie). Etude agrostologique no. 3. Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Mosnier, M. 1967. Les pâturages naturels de la région de Gallayed (Rép. du Sénégal). Etude agrostologique no. 18. Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Naegelé, A.F.G. 1971. Etude et amélioration de la zone pastorale du nord Sénégal. Pâturages et cultures fourragères no. 4. Rome, Italy: Food and Agriculture Organization of the United Nations. Nebout, J.P., and Toutain B. 1978. Etudes sur les arbres fourragers dans la zone sahélienne (Oudalan voltaϊque). Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Palo, R.T. 1985. Chemical defense in birch: inhibition of digestibility in ruminants by phenolic extracts. Oecologia 68:10-14. Pélissier, P. 1979. L'arbre dans les paysages agraires de l'Afrique noire. Pages 37-42 in Le rôle des arbres au Sahel. Canada: International Development Research Centre. Peyre de Fabrègues, B. 1963. Etude des pâturages naturels sahéliens: Ranch du nord-Sanam (Rép. du Niger). Maisons-Alfort, France: Institut d'élevage et de médécine vétérinaire des pays tropicaux. Portères, R. 1957. Un arbre vivant à contre saison en Afrique soudano-zambézienne: Faidherbia albida. Sciences et Nature 19:19-24. Poupon, H. 1980. Structure et dynamique de la strate ligneuse d'une steppe sahéllienne au nord du Senagal. Travaux et Documents no. 115. Paris, France: Institut français de recherche scientifique pour le développement en coopération. Rivière, R. 1977. Manuel d'alimentation des ruminants domestiques en milieu tropical. Paris, France: Ministere de la Coopération. 519 pp. Seignebos, C. 1978. Végétation de clôture et système de défense des végétaux précoloniaux de la zone soudano-sahéllienne (Tchad et Nord Cameroun) Paysage de Parc et Civilisation agraire. Essai de typo-logie. Annals of the University of Chad, Chad. 93 pp. Tanner, J.C. 1988. Acacia fruit supplementation of maize stover diets fed to sheep. M.Sc. Thesis, University of Reading, Reading, UK. 66 pp. Touzeau, J. 1973. Les arbres fourragers de la zone sahéllienne d'Afrique. Thesis, Ecole nationale, Toulouse, France. Trochain, J. 1969. Le rythme phénologique aberrant de Faidherbia albida Del. A. Chev. (Mimosacea). Annales scientifiques de l'Université de Besançon (3)(6):7-13. Van Soest, P.J. 1983. Nutritional ecology of the ruminants. USA: Cornell University Press. 374 pp. Wickens, G.E. 1969. A study of Acacia albida Del. (Mimosoidae). Kew Bulletin 23(2):181-202.
1. Direction de recherche forestiere et hydrobiologique (DRFH)/Institut d'economie rurale (IER), B.P 1704, Bamako, Mali. 2. Centre des recherches zootechniques (CRZ), Sotuba, Bamako, Mali. Cissé, M.I., and Koné, A.R. 1992. The fodder role of Acacia albida Del.: extent of knowledge and prospects for future research. Pages 29-37 in Faidherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey, Niger (Vandenbeldt, R.J., ed.). Patancheru. A.P. 502 324, India: International Crops Research Institute for the Semi-Arid Tropics; and Nairobi, Kenya: International Centre for Research in Agroforestry. |