An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 1 Botany and Distribution Invited Paper The Botany and Distribution of Faidherbia albida P.J.Wood1 Abstract
Introduction Faidherbia albida Del. A. Chev. (syn. Acacia albida) is widely distributed throughout the dry zones of tropical Africa. Although of the Acacieae tribe, it has many unique botanical and phytochemical features, as well as botanical naming, that justify its classification as a separate genus. In western Africa, though not always in eastern, central, or southern Africa, the species has the unique characteristic of shedding its foliage at the start of the rainy season, and of coming into leaf in the dry season. This unexpected "inverted" phenology means that its presence in farmers' fields does not interfere with agriculture, and, indeed, makes it an ideal agroforestry tree for use in combination with crops. It also provides animal fodder in the dry season. These qualities are traditionally well known in rural West Africa, where generations of small farmers have deliberately encouraged it in a kind of cultivated parkland (Pélissier 1980). In southern Africa, mono-specific stands are also thought to result from past cultivation (Fagg and Barnes 1990).
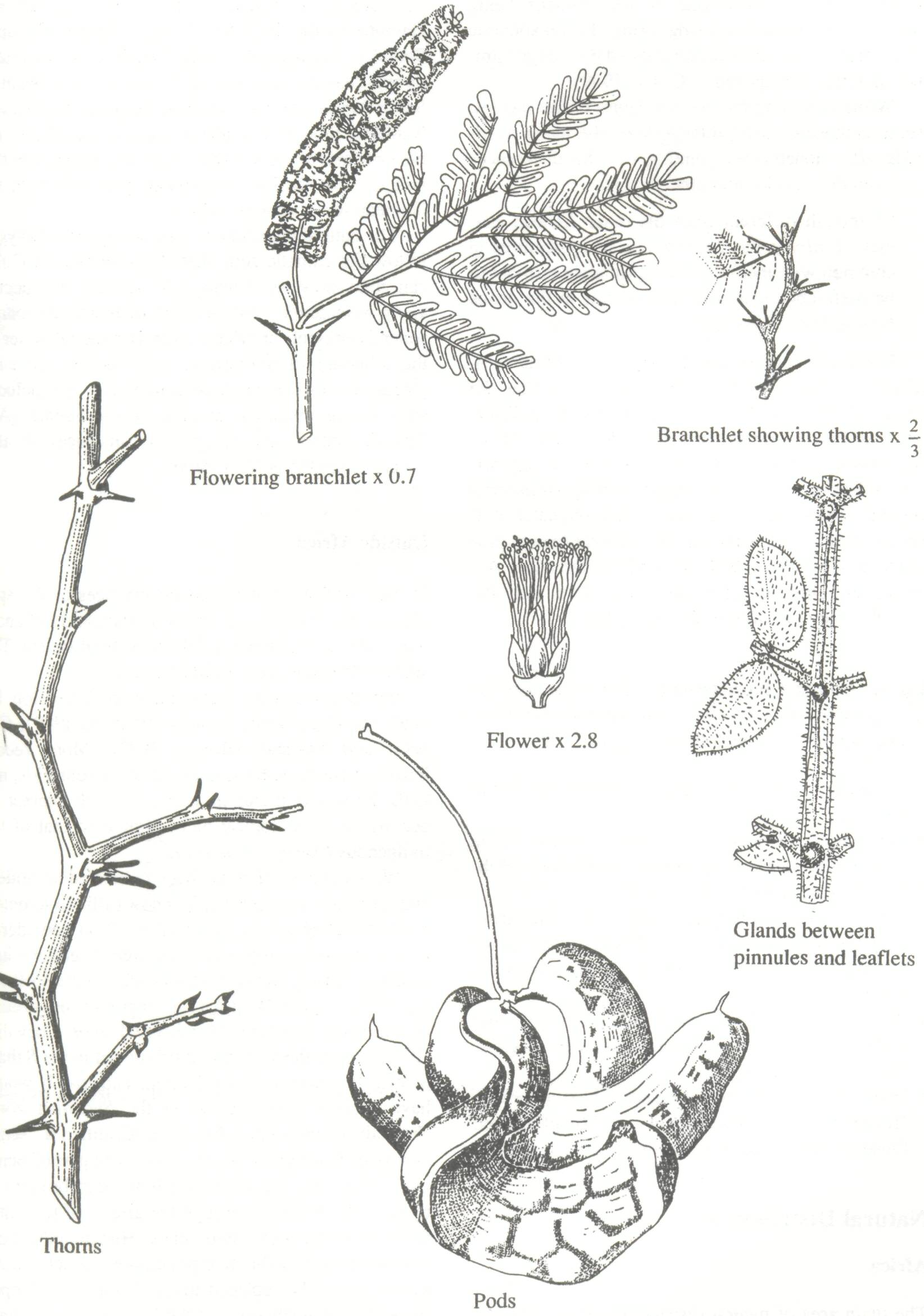
Morphology and General Features Under favorable conditions, F. albida can attain a very large size; heights of over 30 m and a diameter of 1.5 m have been noted along the Cunene River on the Namibia-Angola border (NAS 1975). More usually it reaches 15 to 20 m in height and a breast height diameter of up to 1 m. The crown tends to be shaped like an inverted pyramid in young trees, becoming hemispherical with age. The lifespan is generally about 70 to 90 years, although some individuals of over 150 years are reported from Zambia. The bark of the tree is characteristically dull brown to whitish grey, smooth when young, more fissured and flaky and more cork-like in older specimens. The slash is fibrous, pink to light brown. Botanical Description Leaves. The leaves are typical of the Mimosoideae, compound and bipinnate with leaflets borne along the pinnae. They are highly variable: the petiole varies from 0.5 to 3.7 cm long. The rachis is 3-7.5 cm long and carries 2-12 pairs of pinnae, each of which is 2.5-5.5 cm long and carries 6-23 pairs of leaflets. Leaflets are glabrous to pubescent, bluish green, ob-longate and occasionally submucronate, overlapping slightly, 2.5 to 12 mm in length and 0.7 to 5 mm in width (Fig. 1) (CTFT 1989). Thorns. Thorns occur in pairs at the base of the leaves and are modified, spiny stipules. They are straight and robust, thickened at the base and often (particularly when juvenile) orange or brown at the tip and are 0.2-3.2 cm long. They may be distinguished from those of Acacia species with long thorns, such as A. tortilis subsp raddiana, A. nilotica, or A. seyal, by their basal thickening. Flowers. The flowers are borne in dense axillary panicles 3.5-16 cm long, with a peduncle 2-4 cm long. They appear about 2 months after the tree comes into leaf and are sessile or with a pedicel of up to 2 mm. In color they are successively white, cream, and then yellow, and are strongly perfumed. The calyx is 1-1.7 mm long, glabrous to pubescent, consisting of five sepals. The corolla, 3-3.5 mm long, carries five free petals. The stamens, numbering 40 to 50, are 4-6 mm long and their filaments are fused for about 1 mm. There is also partial fusing between the stamens and the bases of the petals, (epipetaly). The anthers are 0.2-0.4 mm in diameter, lacking glands even in the bud. The ovary is lightly stipitate and bears small hairs. Flowering commences towards the seventh year (Nongonierma 1976; McGahuey 1985). Fruits. The fruit is an indehiscent pod varying from bright orange to reddish brown and 7-9 mm thick, 6-35 cm long, and 1.4-6 cm wide. It falls from the tree about 3 months after flowering. The pod surface is convex on one side and becomes concave on the other, and as the fleshy mesocarp lignifies, it tends to roll into a spiral resembling dried apple peel, hence the common name "Apple Ring Acacia." There is much variation in pod shape both within and between trees. Seeds. Each pod contains 10-29 dark brown shiny seeds, separated by thin septa. They are ovoid, 10 x 6 mm in size and are characterized by an elliptic areolus or hilum measuring about 8x5 mm. The seed coat is tough, waterproof, and leathery, and maintains seed viability for many years.
Identification and Classification The first botanical determination of the species under the name A. albida Del. was made by Delille in 1813 and was based on a specimen obtained in Egypt. Bentham (1875) recognized the species as belonging to the Mimosoideae subfamily of the Leguminosae in the Gummiferae series (Table 1), which is characterized by possessing spiny stipules. Acacia comprises some 1200 species and is the sole genus in the tribe Acacieae, which are characterized by possessing stamens either free or united only at the base.
Because A. albida (F. albida) is quite distinct botanically, having no other species closely related to it, its classification has given rise to a number of taxonomic studies. Baillon (1863) concluded that the staminal filaments were indeed joined at the base, but that this feature was not sufficiently important to justify separating the species from the genus Acacia. On the other hand, Chevalier (1934), using the same distinctive characteristic as well as other features (which are discussed further below), preferred to exclude the species from the genus Acacia and place it in the new monospecific genus Faidherbia. This move established the link between the tribes Acacieae and Ingeae, the latter being another tribe of the subfamily Mimosoideae characterized by the stamens being more or less fused into a single ring. This taxonomic classification has since been followed by a large number of authorities quoted in CTFT (1989).
"Although different from the other African acacias, A. albida has nevertheless many features in common with them. It could ultimately turn out to be preferable to transfer the species to the genus Faidherbia" (Ross 1966). Hutchinson and Dalziel (1958) retain the name A. albida. However, Brain (1987) points out that the chemical constituents of the species are quite different from those of other acacias, and Robbertse (1974) has shown that the pollen also differs. Some authorities still have reservations about keeping Faidherbia separate from the genus Acacia but on balance it seems logical to put the species into the genus proposed by Chevalier (1934), which links the tribes Acacieae and Ingeae. This paper therefore uses the specific epithet F. albida (Del.) A. Chev. Distinguishing Characteristics. The characteristics distinguishing this species from other Gummiferae may be summarized as follows:
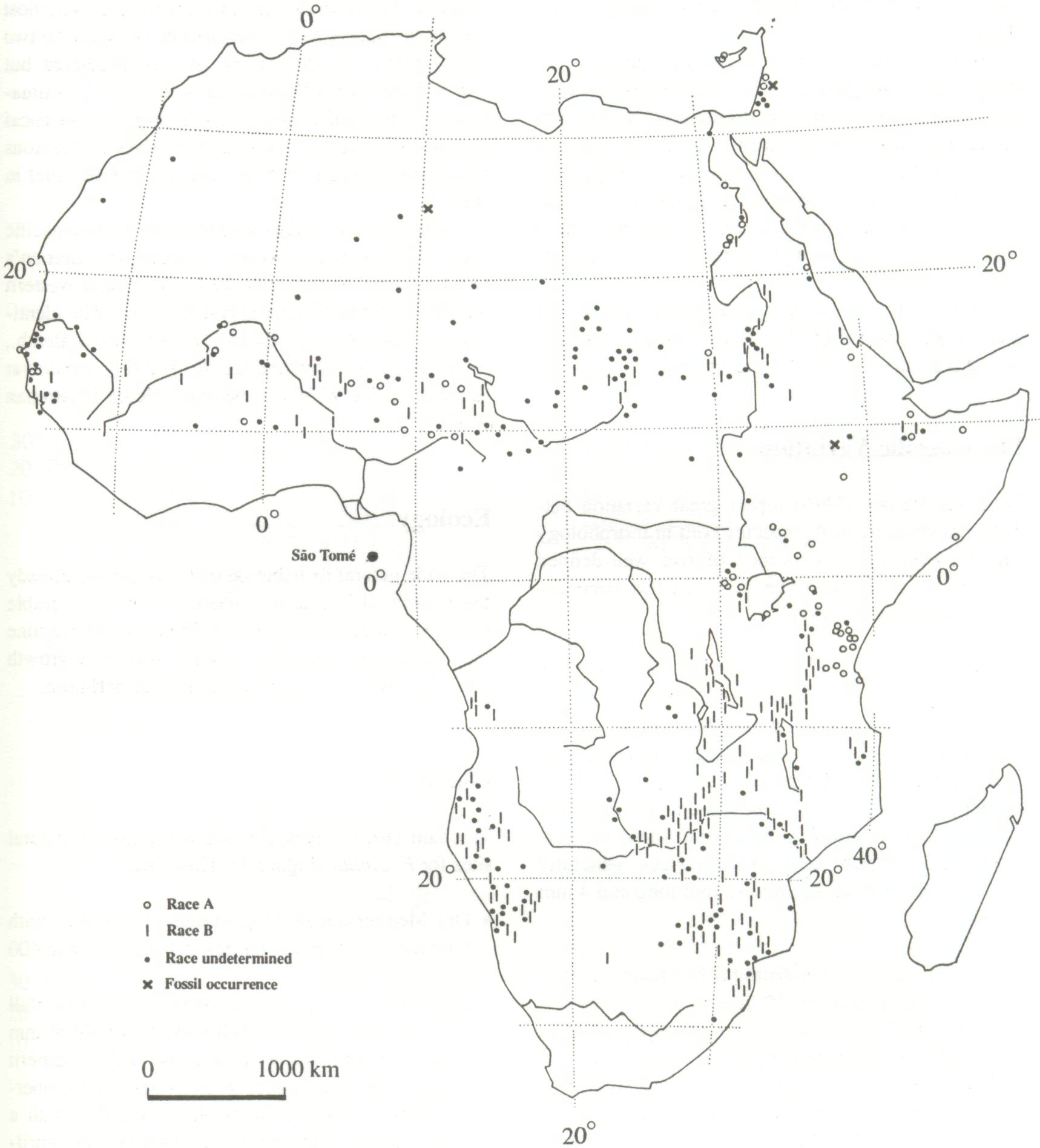
Natural Distribution Africa The main area of natural distribution of F. albida is Africa, as shown in Figure 2. The species occurs right across the African continent from Senegal and Gambia to the Red Sea (Egypt, Sudan, Ethiopia, Somalia, and Kenya). Further south, it is distributed through eastern and central Africa as far as Natal in South Africa, and westwards to Lesotho, Angola, and Namibia, though it is absent from a corridor in the eastern parts of the last two countries. Apart from this area, its distribution corresponds precisely with the Guinean forest zone of Africa. The natural distribution does not appear always to follow the climatic zone that characterizes the Guinean zone, however, because the species also occurs along watercourses and in other areas where ground-water is present, or where there is residual water in the alluvium of a seasonal water course. In such places, the distribution is discontinuous, and includes sites in the mountain massifs of the Sahara (Air, Tibesti, Ennedi, and Hoggar, for instance). It also occurs along the Nile in Egypt. Outside Africa Outside Africa, spontaneous occurrences of the species are found in Yemen, Israel, Jordan, and Lebanon, where the northernmost is 30 km north of Beirut. The species does not occur in Madagascar. Introductions have been made to Ascension Island, the Cape Verde Islands (Wickens 1969), Cy-prus, and Pakistan (Brenan 1983). More recent introductions have been made to other countries, notably Peru and India; in India, it is of interest as occupying an ecological niche similar to that of the indigenous Prosopis cineraria. With a species that has been cultivated so widely and for such a long period, it is now difficult to determine its actual origin. Aubréville (1937) considered that it was originally a riverine tree of eastern and southern Africa, and was introduced into the northern part of its range in West Africa, where it is only found on cultivated or previously cultivated lands. Chevalier (1934) was of the view that it originated in the Sahara before the current desertification, and that it would have become domesticated in the Sudanian zone. Trochain (1969) claimed to have identified a "semi-climax of F.albida in the south-western part of Senegal," where the species would have been brought in by nomadic peoples (through the effects of their animals) in particularly hard times. Human influences thus developed "stable tree populations which, on the sandy clay soils, replaced the different natural open forest climax formations of the Sudanian zone, these being Combretiun glutinosum on the lighter, sandier soils and Acacia seyal on the heavier sandy clays" (Felker 1978).
On the other hand, the two separate races of the species are regarded by Wickens (1969) as an indication of a northern origin for the species.
Fagg and Barnes (1990) report great variation between provenances of the species both in morphology and phenology. The existence of two well-defined races of F. albida in eastern Africa has been described by Brenan (1959):
The two races are not distinguished taxonomically because a large number of intermediate forms are encountered in certain parts of the range. Ross (1966) states that the characteristics of the two races described by Brenan are not strongly correlated and, as Brenan points out, they appear more as heterogeneous features to the extent that intermediate forms show peculiarities of both races. However, there is confusion only in some regions. Ross indicates that most specimens from the southern part of the range fall within Race B, which is the only one represented in Lesotho, Namibia, Zambia, Zimbabwe, Mozambique, Malawi, and most of Tanzania. Race A is found in northern and eastern Tanzania, Kenya, Uganda, Somalia, and Ethiopia. In Sudan and in West Africa, Race B is common, but many specimens combine characteristics of both races; i.e., large leaves and glabrousness or small leaves and pubescence. In Egypt and in the northern populations, most specimens appear to be intermediate between the two races, being generally more or less pubescent but with smaller leaves than normal in Race B. The situation is probably the same in the Yemen, but botanical material is currently inadequate. In Cyprus, glabrous leaves predominate, but they are larger than usual in Race A. Nongonierma (1976) identified four intraspecific taxa, which do not, however, compare with Brenan's races, and are based only on northern and western African material (Fagg and Barnes 1990). The identified varieties (Acacia albida var senegalensis Benth., A. albida var microfoliata De Wild, and A. albida var variofoliata De Wild.) all belong to Race B (Brenan 1983).
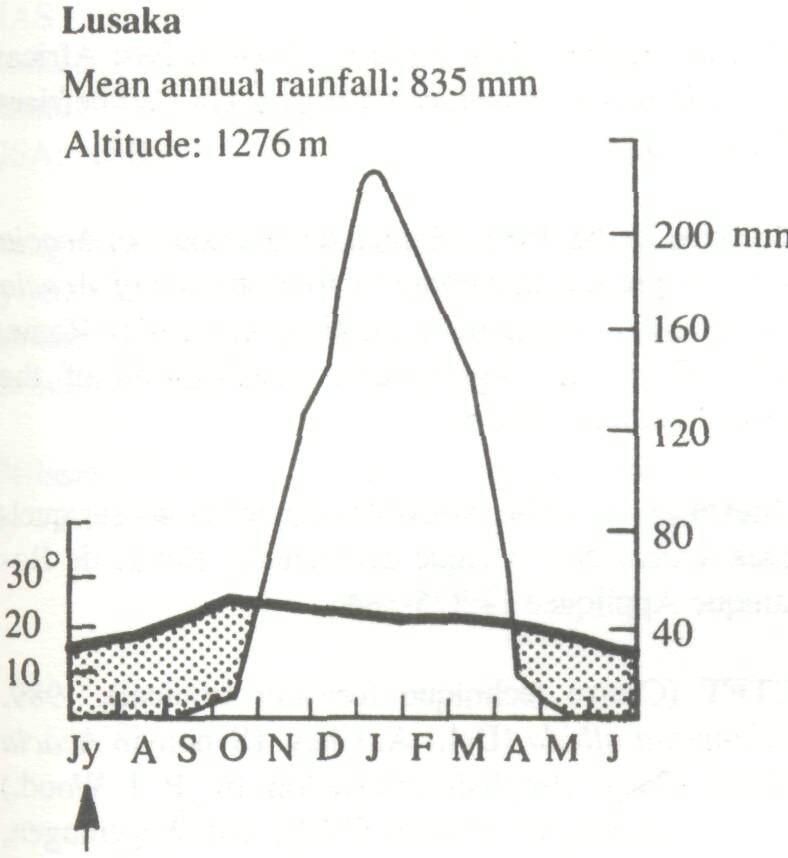
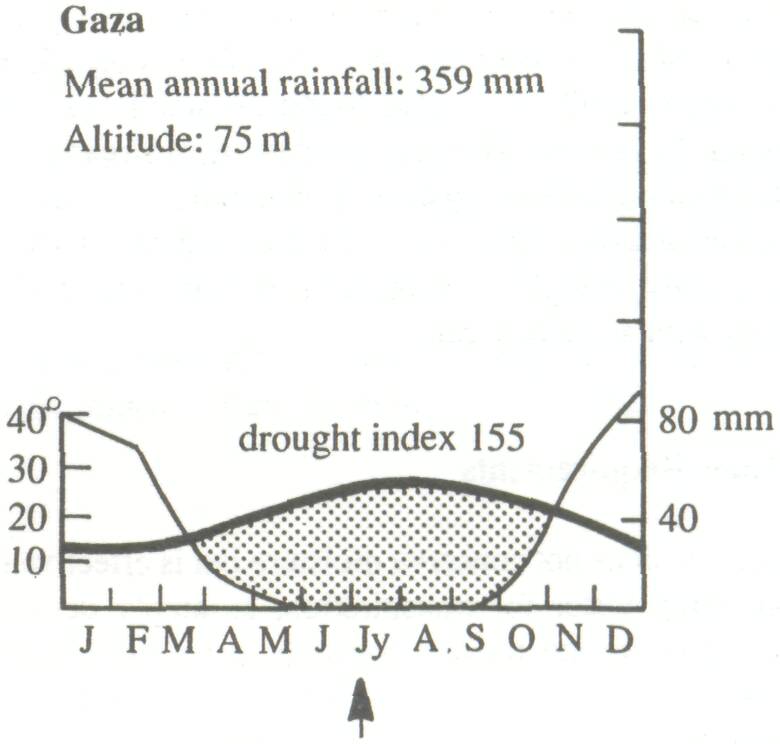
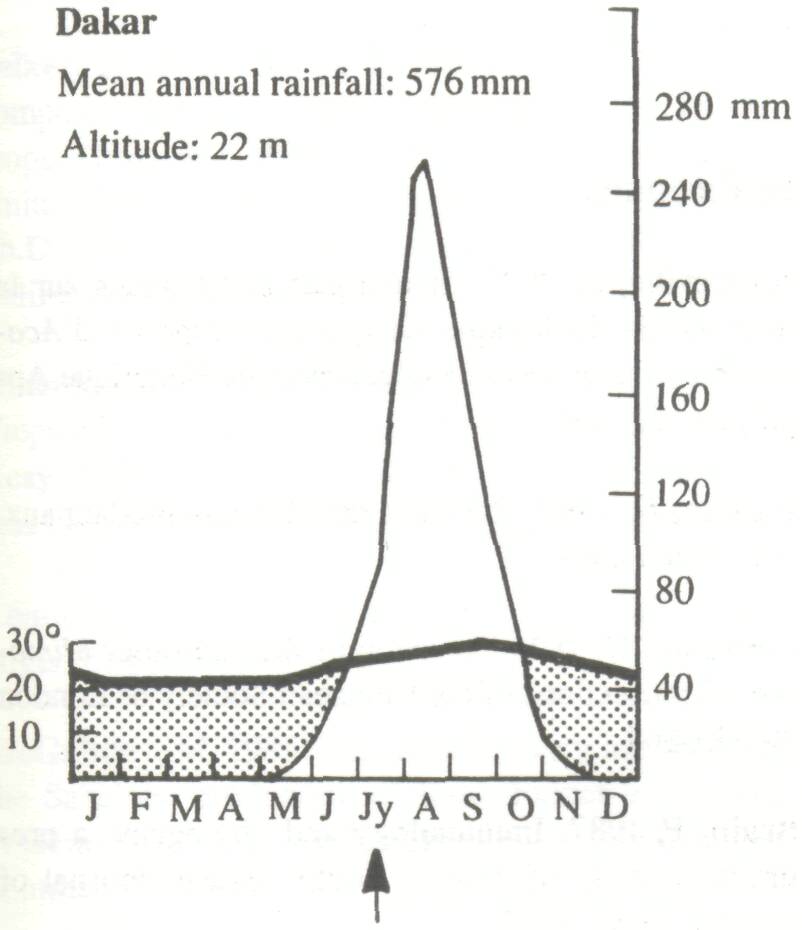
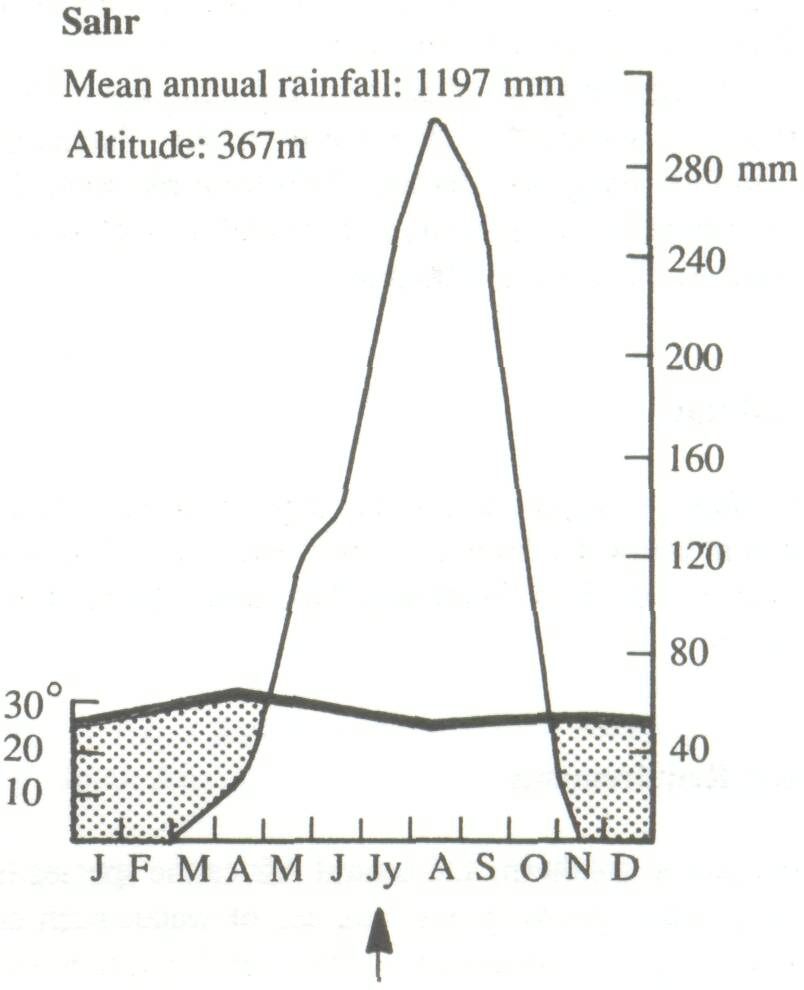
The wide natural distribution of F. albida has already been noted. It is clearly a species with considerable ecological adaptability and it is important to examine the relationship between climate, soil, and growth and, if possible, to define an ecological optimum. Climate Trochain (1969) gives climatic diagrams for natural sites for F. albida (Figure 3). These are:
These climatic diagrams illustrate the wide range of climatic conditions in which the species will grow, the only constant feature being a long and well-defined dry season. Saturation deficit, particularly in the Sudan and Sahelian Zones, is an important parameter, not often included in climatic records. Obviously the species will stand very low air humidities, as in the mountains of the Sahara. In some countries, however, the air humidity in the dry season is often relatively high, as in eastern Africa. In the eastern Mediterranean, the species survives frost.
Altitude F. albida will grow at a wide range of altitudes, from 270 m below sea level near the Dead Sea in Palestine to over 2000 m in Ethiopia and the Jebel Marra of the Sudan. Soil Requirements In eastern, southern, and central Africa the species is often found growing near sources of water, such as along rivers, on the shores of lakes, and in gullies and ravines. In all these situations, the soils tend to be alluvial or hydromorphic. These seem to be the ideal sites; in Sudan, for instance, the greatest concentrations of the species occur in wide belts along the sandy alluvia of the drainage basin of the Jebel Marra (Wickens 1969). It is also found on stabilized sand dunes. In western Africa, as we have seen already, the distribution of the species is generally a result of human activity. Here, it prefers deep, lighter sandy or silty soils, though occasionally it is found on lateritic soils with a shallow pan. Water Requirements The tree does not conserve moisture but is effective in obtaining water for transpiration, as amply demonstrated by its vegetative vigor in the dry season, in the total absence of rain, when temperatures are high, humidity is low, and evapotranspiration is at a maximum. To achieve this, its root system has a taproot that grows very rapidly and will reach the water table if it is within range; depths of about 40 m have been recorded (Lemaitre 1954). When on its preferred sites, notably in association with water, the species prefers deep sandy soils easily exploited by its root system. Acknowledgment. The bulk of this paper relies heavily on the standard monograph on the species produced by the Centre technique forestier tropical (CTFT 1989), to which grateful acknowledgment is made.
Aubreville, A. 1937. Remarques écologiques sur la distribution écologique de quelques espèces d'Aca-cias en Afrique Occidentale. Revue de Botanique Appliquée. 796 pp. Baillon, H. 1863. Révision des Acacias médicinaux. Adansonia 4:85-127. Bentham, G. 1875. Revision of the sub-order Mimoseae. Transactions of the Linnaean Society of London 30:335-664. Brain, P. 1987. Immunology and phylogeny: a preliminary study of Acacia. South African Journal of Science 83:422-427. Brenan, J.P.M. 1959. Flora of Tropical East Africa: Leguminosae Mimosoideae. London, UK: Whitefriars Press. 173 pp. Brenan, J.P.M. 1983. Manual on taxonomy of Acacia species: present taxonomy of four species of Acacia (A. albida, A. Senegal, A. nilotica, A. tortilis). Rome, Italy: Food and Agricultural Organization of the United Nations. 47 pp. Chevalier, A. 1934. Nouvelles observations sur quelques Acacia de l'Afrique occidentale. Revue de Botanique Appliquee 14:875-884. CTFT (Centre technique forestier tropical). 1989. Faidherbia albida (Del.) A. Chev. (Synonym Acacia albida Del.). (English translation by P J Wood.) Nogent-sur-Marne, France: CTFT, and Wageningen, Netherlands: Centre technique de cooperation agri-cole et rurale. 72 pp. Fagg, C.W., and Barnes, R.D. 1990. African Acacias: Study and aquisition of the genetic resources. Final Report on Overseas Development Administration Research Scheme R.4348. UK: Oxford Forestry Institute. 170 pp. Fahn, A., Werker, E., and Boas, P. 1986. Wood anatomy and identification of trees and shrubs of Israel and adjacent regions. Jerusalem, Israel Academy of Sciences and Humanities. 221 pp. Felker, P. 1978. State of the art: Acacia albida as a complementary permanent intercrop with annual crops. Grant no. AID/afr-C-1361. Washington, D.C., United States Agency for International Development. Ph.D. thesis, University of California, Riverside, California, USA. 133 pp. Hutchinson, J., and Dalziel, J.M. 1958. Flora of West Tropical Africa, Vol. 1 Part II. 2nd edn. (Revised by Keay, R W J.). London, UK: Crown Agents for Overseas Governments and Administrations. 499 pp. Lemaître, C. 1954. Le Faidherbia albida. Thesis, Niger. 62 pp. McGahuey, M. 1985. Impact of forestry initiatives in the Sahel on production of food, fodder and wood. Washington, D.C., USA: Chemonics International. (Limited distribution.) NAS (National Academy of Sciences). 1975. Under-exploited tropical plants with promising economic value. Report of an Ad Hoc Panel. Washington, D.C., USA: NAS. Nongonierma, A. 1976. Contribution à l'étude biosystématique du genre Acacia Miller en Afrique occidentale. II. Caractères des inflorescences et des fleurs. Bulletin IFAN 38:487-642. Pélissier, P. 1980. L'arbre dans les paysages agraires de 1'Afrique noire. L'arbre en Afrique tropicale: la fonction et le signe. Cahiers ORSTOM, Série Sciences Humaines 17(3-4):131-136. Robbertse, P.J. 1974. A scanning electron microscope investigation of the pollen of South African Acacia species. Journal of South African Botany 40(2):91-99. Robbertse, P.J., Venter, G., and van Rensburg, J.H. 1980. The wood anatomy of the South African acacias. IAWA Bulletin 1(3):93-103. Ross, J.H. 1966. Acacia albida Del. in Africa. Boletin de Sociedade Broteriana 2(40):187-205. Ross, J.H. 1979. A conspectus of the African Acacia species. Pages 83-85 in Memoires of Botanical Survey, South Africa. Ross J.H. 1981. An analysis of the African Acacia albida species: their distribution, possible origins and relationships. Bothalia 13(3-4):389-413. Trochain, J.L. 1969. Le rythme phénologique aberrant de Faidherbia albida (Del.) A. Chev. (Mimosaceae). Annales scientifiques de l'Université de Besançon, 3(6):7-13. Vassal, J. 1979. Intérisêt de l'ontongénie foliaire pour la taxonomie et la phylogénie du genre Acacia. Bulletin de la Societé botanique française 3:55-65. Wickens, G.E. 1969. A study of Acacia albida Del. (Mimosoideae). Kew Bulletin 23(2):181-202.
I. Overseas Development Administration, London, UK. Wood, P.J. 1992. The botany and distribution of Faidherbia albida. Pages 9-17 in Faidherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey, Niger (Vandenbeldt. R.J., ed.). Patancheru, A.P. 502 324, India: International Crops Research Institute for the Semi-Arid Tropics; and Nairobi, Kenya: International Centre for Research in Agroforestry. |

 g
g