An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 4 Site Effects, Silviculture, and Symbiosis Session Papers Symbiosis Presence of Bradyrhizobia under Acacia albida N. Dupuy and B. Dreyfus1 Abstract
Introduction It is thought that soil improvement beneath Acacia albida is linked to its nitrogen-fixing ability. Abundant nodulation on roots of A. albida trees growing in rice fields of Casamance, Senegal (annual rainfall 1000 mm) has been observed. However, no nodules have been found on roots of adult trees in the Sahelian zone (annual rainfall <1000 mm). Thus, there is a question whether A. albida does indeed fix nitrogen in arid zones. In order to answer this, two approaches were considered. The first consisted of determining the presence of nodules on root systems as well as the existence of specific Rhizobium populations in the rhizosphere. The second approach involves measuring nitrogen fixed by tree-host associations using the isotope labelling method. Studies using the first approach are presented here. Two holes were drilled beneath adult A. albida in order to sample the distribution of specific Rhizobium populations throughout the soil profile. Subsequently, Rhizobium strains from upper and lower horizons were collected and their nitrogen-fixing capability determined.
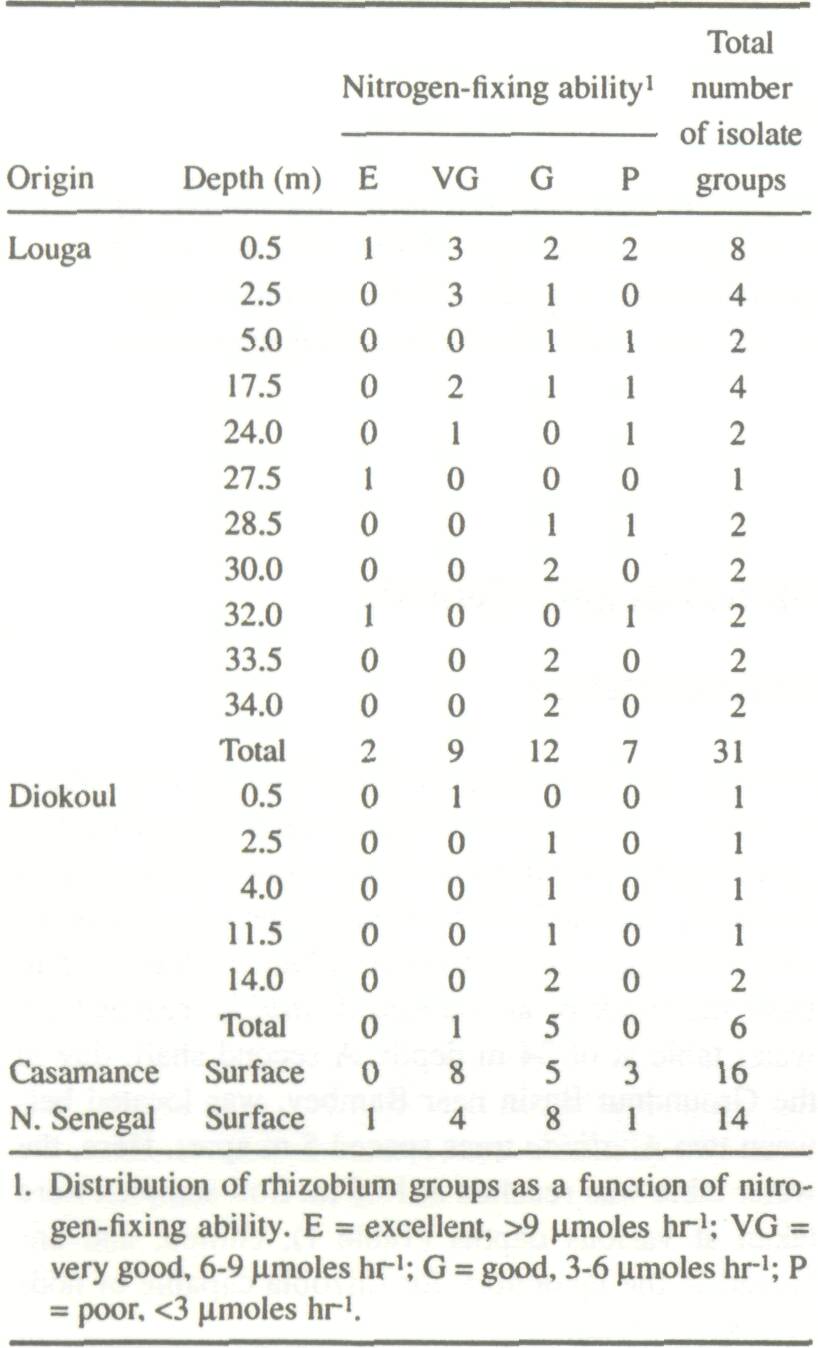
Sampling Method Borings were done using a light, hand-operated auger to which 1.5-m long aluminum shafts (Dormer's Engineering, South Murwillumbah, NSW 2484, Australia) were attached as the hole deepened. One shaft sunk in Louga (200 km north of Dakar), located 8 m from the trunk of an isolated A. albida, reached the water table at of 34 m depth. A second shaft, dug in the Groundnut Basin near Bambey, was located between two A. albida trees spaced 5 m apart. Here, the water table was reached at 16.5 m. Soil samples were taken at various depths (Table 1), chilled, and analyzed in the laboratory for rhizobia capable of nodulating A. albida. Rhizobium Densities in Soil Samples Rhizobium densities of the soil samples were estimated using the most probable number method (Brockwell 1980). A. albida seeds were sterilized in concentrated sulfuric acid for 1 h, then washed several times in sterile water and finally germinated in petri dishes on agar-agar water (0.8%) for 48 h. Roots of the germinated seedlings were placed in test tubes containing Jensen medium; the shoots remained exposed to air. The seedlings were placed in a culture chamber and inoculated after 5 days with 1 mL of a suspended soil dilution. For each dilution, tubes showing nodulation after 4 weeks were counted. Non-inoculated controls did not nodulate.
Nodules from inoculated tubes were sterilized in a 0.1% HgCl2 solution for 3 min, washed in distilled water 6-7 times, then ground in a drop of water with a sterile glass rod. Strains of Rhizobium were isolated on a YMA medium following the procedure of Vincent (1970). Evaluation of Nitrogen-Fixing Capability After removing the aerial parts, the liquid culture medium was discarded and the tubes were closed with stoppers. Acetylene was injected (10% of tube volume) into the tube, and acetylene reduction was measured by chromatography in the gaseous phase using the method of Hardy et al. (1968) after a 30-min incubation period at 28°C. Rhizobium activity remained linear for 90 min. The strains were divided into four classes according to the acetylene reduction assay results (Table 1).
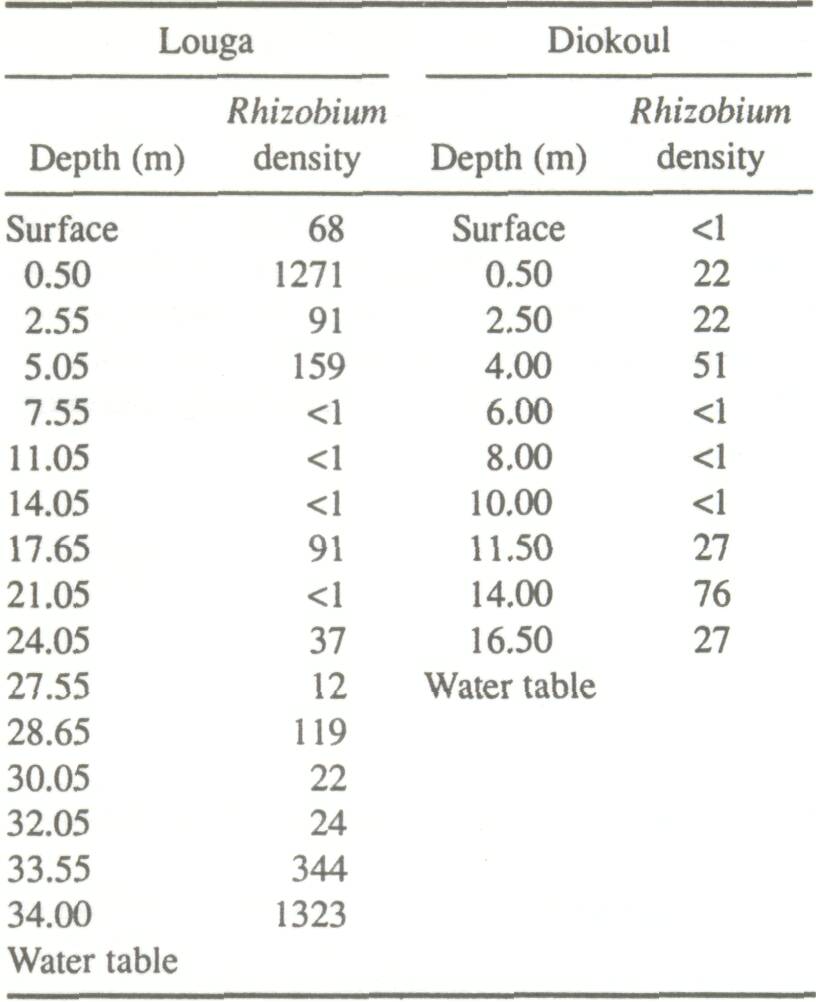
Rhizobium Distribution as a Function of Depth Contrary to expectation, Rhizobium population densities were highest (Table 2) at the greatest depth measured (Louga). Also, certain intermediary zones (7-14 m at Louga, 6-10 m at Diokoul) contained less than 1 rhizobia per gram of soil throughout the profiles studied. Rhizobium populations were less numerous at the Diokoul site than the Louga site. This is probably due to differences in soil texturethe Diokoul soil is much more clayey and therefore probably not as favorable to Rhizobium multiplication and development as are the sandy dune soils of Louga.
Of the 67 strains isolated, all except two (ORS167, ORS173) demonstrated slow growth on the YMA medium and were thus considered as Bradyrhizobium. Eleven of the 31 Louga strains were classed as very good or excellent in terms of nitrogen-fixing capability. Two of the excellent strains (ORS130 and ORS136) were isolated at depths of 27.5 and 32 m. At Diokoul, however, no strains classed as very good or excellent were isolated below 0.5 m.
Until now, it has been believed that Rhizobium populations reside only in the upper horizons of the soil; i.e., at depths not exceeding 1-2 m (Alexander 1977). Before this experiment, only the research team led by Virginia et al. (1986), following the work of Felker and Clark (1982), studied Rhizobium distribution in the soil. Their results showed that Rhizobium populations beneath Prosopis glandulosa in California could attain densities of 10 x 103 g-1 of soil, at depths reaching 4-6 m. The present study has demonstrated that Rhizobium can heavily populate soils (up to 1.3 x 103 per g of soil) beneath A. albida at depths never previously thought to have been inhabited by this bacterium. Jenkins et al. (1987) showed that two distinct Rhizobium populations existed under P. glandulosa, according to depth. In the upper horizons, Rhizobium predominates, whereas deeper in the soil, Bradyrhizobium prevails. With the exception of two strains, all of the Rhizobium strains isolated in this experiment exhibited traits similar to those of Bradyrhizobium (Jordan 1984). It is possible that the lower-horizon strains and those of the upper horizons form two distinct subgroups. Since environmental conditions at greater depths are relatively constant, the lower-horizon Bradyrhizobium may differ from upper-horizon strains which are better adapted to wide environmental fluctuations. A taxonomic study based on the comparison of total protein profiles and of nutritional traits is under way to verify this hypothesis. The presence of nodules on shallow roots of naturally occurring A. albida in the Sahelian zone is rare. This absence of nodules may result from the inhibitive effect of limiting factors of the environment, notably drought, and not genetic determinants linked to the plant. A. albida nodulates abundantly on continually moist soil, as in the rice fields at Casamance. The absence of large Rhizobium populations at greater depths at Casamance may have been due to a lack of oxygen or the difficulty of observing roots. Isotopic tracing may prove useful in determining whether A. albida fixes nitrogen in the Sahelian zone. Lower-horizon strains which proved to be very effective nitrogen-fixers could be used to colonize deep root systems of A. albida.
Alexander, M. 1977. Introduction to soil microbiology. 2nd ed. New York, USA: J. Wiley & Sons. Pages 21-24. Brockwell, J. 1980. Experiments with crop and pasture legumes: Principles and practice. Pages 417-488 in Methods for Evaluating Biological Nitrogen Fixation (Bergensen, F.J., ed.). New York, USA: John Wiley & Sons Ltd. Felker, P., and Clark, P.R. 1982. Nodulation and nitrogen fixation in 3 m-long phraetophytically simulated soils columns. Plant and Soil 61:297-305. Hardy, R.W., Holsten, R.D., Jackson, E.K., and Burns, R.C. 1968. The acetylene-ethylene assay for N2-fixation: Laboratory and field evaluation. Plant Physiology 43:118-120. Jenkins, M.B., Virginia, R.A., and Jarrell, W.M. 1987. Rhizobial ecology of the woody legume mes-quite (Prosopis glandulosa) in the Sonoran Desert. Applied Environmental Microbiology 53:36-40. Jordan, D.C. 1984. Family III. Rhizobiaceae Conn 1938. Pages 234-244 in Manual of systematic bacteriology, vol.1 (Krieg, N.R., and Holt, J.G., eds.). Baltimore, UK: Williams and Wilkins Co. Vincent, J.M. 1970. A manual for practical study of root-nodule bacteria. IPB Handbook. Oxford, UK: Blackwell Scientific Publications. Virginia, R.A., Jenkins, M.B., and Jarrell, W.M. 1986. Depth of root symbiont occurrence in soil. Biology and Fertily of Soils 2:127-130.
1 Departement de microbiologie, Institut francais de recherche scientifique pour le developpement en cooperation (ORSTOM), B.P. 1386, Dakar, Senegal. Dupuy, N., and Dreyfus, B. 1992. Presence of Bradyrhizobia under Acacia albida. Pages 145-148 in Faidherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey, Niger (Vandenbeldt, R.J., ed.). Patancheru, A.P. 502 324, India: International Crops Research Institute for the Semi-Arid Tropics; and Nairobi, Kenya: International Centre for Research in Agroforestry. |