An e-publication by the World Agroforestry Centre
FAIDHERBIA ALBIDA
in the West African
Semi-Arid Tropics

|
|
An e-publication by the World Agroforestry Centre |
|
FAIDHERBIA ALBIDA |
|
|
Session 4 Site Effects, Silviculture, and Symbiosis Session Papers Symbiosis Endomycorrhiza Infection in Young Faidherbia albida: Influence on Growth and Development M. Ducousso1 and J.P. Colonna2 Abstract
Introduction Faidherbia albida, like most of the Acaceae, is capable of simultaneously forming nitrogen-fixing nodules and vesicular arbuscular endomycorrhizae (VAM) on its roots (Ducousso 1990). These infections have been shown to improve seedling development, and have been the focus of studies under controlled conditions in the laboratory and the nursery. Efforts to successfully extend these beneficial effects to transplanted seedlings have failed. Outplantings of Acacia holosericea, Acacia Senegal, and Acacia raddiana in Senegal demonstrated that beneficial effects of endomycorrhizal inoculation diminished after 1 or 2 years. Interest in F. albida as an element of Sahelian agroforestry systems (CTFT 1988) has stimulated research on mycorrhizal infection on this species in the field and its effect on growth. Two experiments were simultaneously carried out on nonsterilized soil representative of F. albida park zones in Senegal. In the first experiment, we monitored the effects of seedling inoculation in polyethylene bags (vol = 1 L), using standard nursery techniques. In the second experiment, seedlings were cultivated in PVC tubes (25 cm in diameter) of various lengths. This experiment represented an intermediate step between the nursery conditions (i.e., bags) and field conditions by allow-ing observation of root systems and mycorrhizal development on plants growing in a larger volume of soil.
F. albida seeds (provenance 90/2709, DRPF/ISRA, Dakar, Senegal) were pretreated with concentrated sulfuric acid for 1 h, rinsed well with tap water and then germinated for 48 h on moist sand. After germination, seed coats were removed and the seedlings were planted in the nursery in either polyethylene bags (vol=1 L) or in 0.5-m (vol=25 L), 1.0-m (vol=50 L), or 1.5-m (vol=75 L) PVC tubex (25 cm in diameter) (Fig. 1). All pots were filled with sifted Deck soil (Table 1). Plants were watered daily throughout the duration of the experiments.
Table 1. Principal physico-chemical characteristics of Deck soil, Bambey, Senegal, 1990
Experimental Design The first experiment had two factorsinoculation (a noninoculated control, C, and inoculation with Glomus mosseae, G), and sampling date (1, 2, 3, 4, 6, 8, 10, and 12 weeks). Each treatment was replicated three times in a completely randomized design. The second experiment involved the same factors, but sampling commenced when the first roots appeared at the bottom of each of the three tube sizes (0.5, 1.0, and 1.5 m in length). There were five replications of each treatment in a completely randomized design. Main stem height, dry above-ground biomass, root length, and number of nodules were measured for both experiments. The percentage of roots infected with endomycorrhiza was estimated with a l0 x mi-croscope after staining with Trypan blue in lactophenol (Phillips and Hayman 1970). In the first experiment, this was done using 100 pieces of root fragments, each 1 cm long, chosen randomly from the root system of each plant. For very young seedlings, samples were taken from the bulked root systems of all sampled plants. In the second experiment, all roots present at the following depths were observed:
Plants were harvested after 7 weeks for the 0.5-m PVC tubes, 9 weeks for the 1.0-m PVC tubes, and 12 weeks for the 1.5-m PVC tubes.
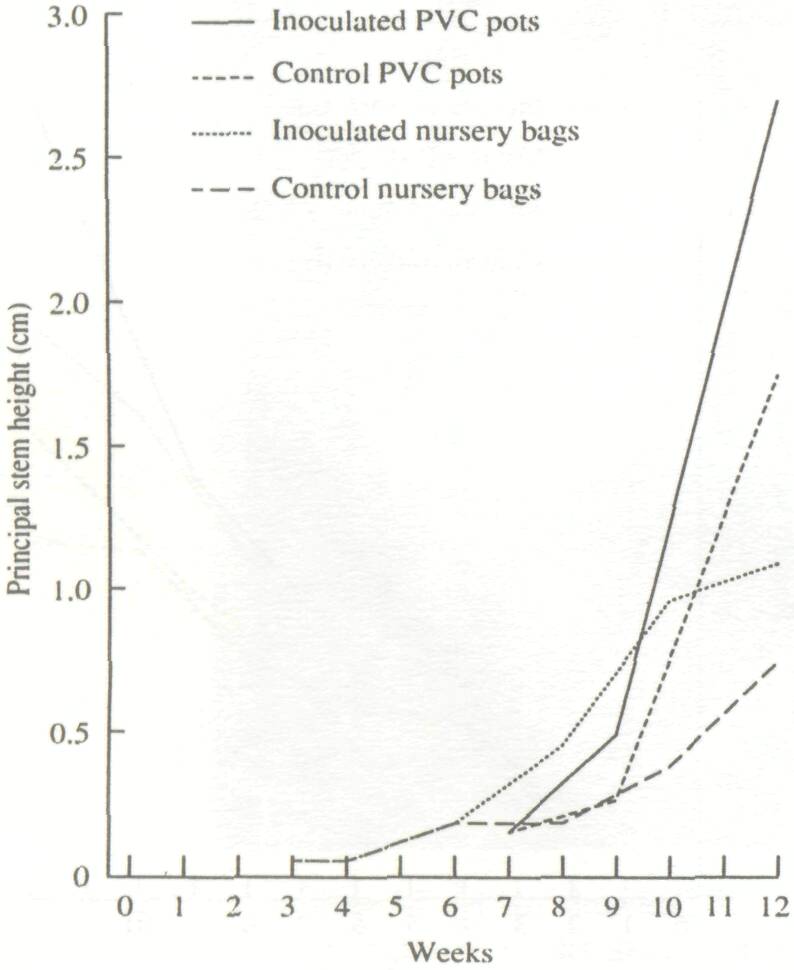
Inoculation by Glomus mosseae had a clear positive impact on plant height. After 12 weeks, this effect was even more noticeable on plants grown in PVC tubes (68% height increase over the control) than on plants grown in polyethylene bags (34%, Fig. 2). Likewise, above-ground biomass of inoculated plants grown in polyethylene bags exceeded the control by 82%. Plants grown in PVC tubes produced 112% more above-ground biomass than the noninoculated control grown in bags. This is because the limited volume of the bags suppressed root growth after the 8th week, whereas the larger PVC tubes allowed unrestricted root development up to the 12th week.
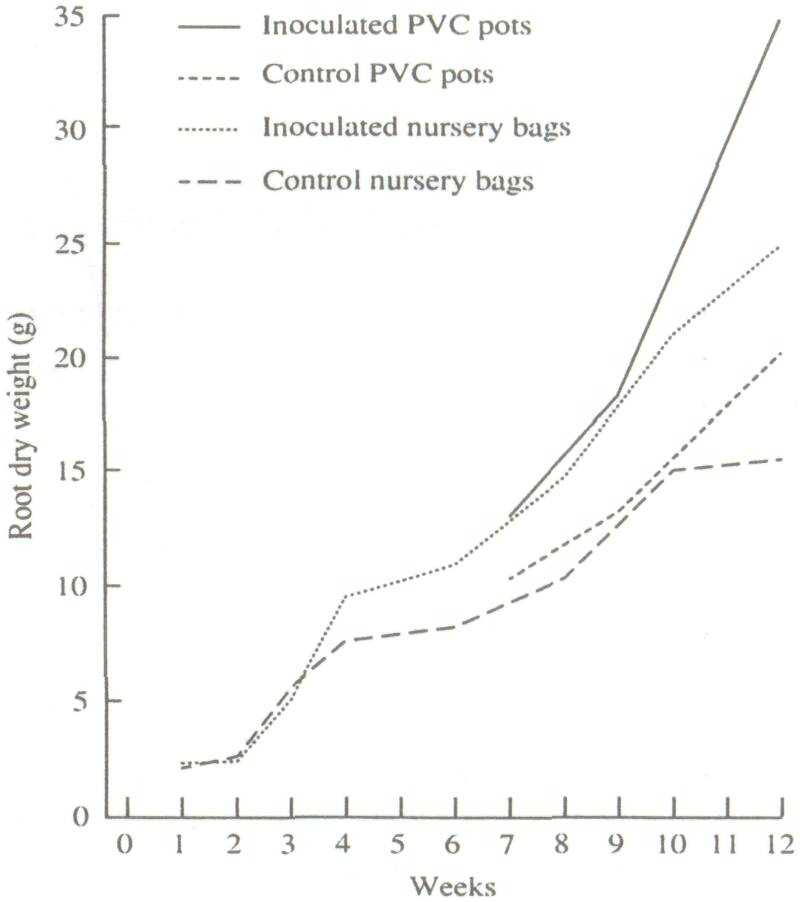
Besides evidence of a slight depressive effect on 1-week-old plants, inoculation by Glomus mosseae had no effect on taproot elongation. Root length in polyethylene bags averaged 32 cm at 12 weeks. At this time, limited growing space caused the roots to coil up at the bottom. In the PVC tubes, where taproot growth was unhindered, root lengths averaged 47 cm in 7 weeks, 95 cm in 9 weeks, and 143 cm in 12 weeks. Inoculated plants in polyethylene bags produced 44% more root biomass than the control, whereas those in PVC tubes yielded 57% more root biomass than the control (Fig. 3). This difference is strongly linked to the large increase in fibrous roots observed on the inoculated plants. This increase evidently resulted in increased mineral absorption capacity which led to growth variations among the treatments. With the exception of plant height, other growth parameters measured were greater than those recorded by Gupta et al. (1973) at equivalent ages. This could be due to differences in growth conditions, growth period, and/or genotype.
Inoculation at the time of outplanting caused necrosis of the taproot on 20% of the plants. Plants thus affected developed 2-6 secondary roots. Inoculating the plants 2-3 weeks after outplanting may have two advantages, i.e., a well-developed root system would ensure a good infection, and the well-formed taproot would not be injured. Twelve weeks is the minimum time required in the nursery for F. albida. These data show that, in this time, the average seedling grown in 1-L pots would have a 32-cm taproot and a root biomass of 0.75 g for the noninoculated control or 1.08 g for inoculated plants. To reduce or eliminate production time in the nursery and to take advantage of the growth potentials demonstrated by these experiments, use of larger containers, or even direct seeding, is a preferable alternative. Nodule counts were taken after the second week, and correlated positively with root development. Inoculation by Glomus mosseae greatly enhanced spontaneous nodulation by Bradyrhizobium (Dreyfus and Dommergues 1981) already present in the soil. Colonna et al. (1990a and 1990b) and Ducousso (1990) suggested that this effect is due to better phosphorous nutrition resulting from infection with endomycorrhiza. Soil volume had a positive effect on plant development, but the degree of development depended on whether the plants were inoculated. For inoculated plants, the quantity of available phosphorus and Bradyrhizobium already in the soil (which would increase with increasing soil volumes), affected the degree of nodulation in the larger PVC tubes. On noninoculated plants, however, seedlings grown in larger volumes of soil had fewer nodules than plants grown in 1-L bags. This could be because nitrogen availability was sufficient in the PVC tubes but limited in the bags. For the noninoculated plants produced in bags, the rate of contamination by local endomycorrhizal fungi: varied with plant age, i.e., 0% for individuals at 1, 2, 3, and 4 weeks, and 40% (maximum) for 12-week-old plants. No endomycorrhiza was observed on non-inoculated plants grown in PVC tubes. Since the soil used was not sterilized at the beginning of the experiment, the appearance of endomycorrhizal contaminants would be expected. The absence of infection of plants raised in PVC tubes remains unexplained. After 6 weeks, 90% of plants raised in bags showed signs of endomycorrhizal infection. One week after inoculation, the first appressoria and intra-root hyphae were noticeable. The first arbuscules were visible in the second week of culture. The rate of infection increased steadily until the 6th week of culture, and then stabilized between 79-90%. In the PVC tubes, infection rate decreased with depth. No endomycorrhiza was observed below 70-80 cm. Endomycorrhizal infection was profuse at 0-10 cm where it was 76% at week 7, 82% at week 9, and 95% at week 12. At 20-30 cm, a more moderate endomycorrhizal development occurred6% at week 7,12% at week 9, and 41% at week 12. Virtually all of the roots infected by Glomus mosseae were located in the upper part of the plant root system. It is quite possible that this temperate variety of Glomus mosseae would not endure extreme variations in surface soil temperature typical in the Sahelian dry season. If an introduced strain happens to survive, its ability to infect new roots at depth would diminish as the root system continued to develop at depth. These results explain, at least in part, the poor results of the inoculation trials carried out in Senegal with Glomus mosseae (Cornet 1982, Cornet et al. 1982, Jaques 1986).
Results of the effects of endomycorrhizal inoculation on growth of F. albida seedlings illustrate the importance of infection in improving the silviculture of this species. Positive effects on growth parameters emerged following inoculation by Glomus mosseae and were enhanced when larger containers were used. We believe that successful manipulation of mycorrhizal infection in F. albida in the field could lead to spectacular results. The failure of endomycorrhizal fungi to develop at greater depths, even under optimal moisture conditions, requires further investigation. As in the case of the inoculation trials undertaken in Senegal, utilization of temperate strains of Glomus mosseae may explain the limited benefits demonstrated here. In future, research should not be limited to the surface horizons only , as is often the case, but also cover the mycorrhizal infection at greater depths. Roots of F. albida in Senegal have been found at depths attaining 34 m (Dupuy and Dreyfus, these proceedings). Collection and conservation of endomycorrhizal fungi of F. albida is important. Also, improved methods for producing sufficient quantities of inoculum for nurseries is essential. Inoculation techniques should emphasize development of controlled infection at depths using selected strains which can successfully compete with wild types already in the soil.
Colonna, J.P., Ducousso, M., and Badji, S. 1990a. Establishment in a sterilized Dior soil of a tripartite symbiosis: Acacia senegal-Rhizobium sp. ORS.1007-Glomus mosseae. Bulletin IGSM 18:105-1 1 1. Colonna, J.P., Ducousso, M., and Badji, S. 1990b. (In press.) Peut-on améliorer le fonctionnement de l'Acacia Senegal (L.) Willd. et du modèle symbiotique "Acacia senegal-Rhizobium" ? Physiologie des arbres et arbustes en zones arides et semi-arides. Nancy, France. Cornet, F. 1982. Recherches préliminaires sur la symbiose de deux Acaceae tropicaux (A. raddiana Savi and A. holosericea A. Cunn.) avec Rhizobium sp et un champignon endomycorrhizien Glomus mosseae Nicol. et Gerd.) DEA Thesis, University of Lyon, Lyon, France. No. 1, 58 pp. Cornet, F., Dieni, H.G., and Dommergues, Y.R. 1982. Effet de l'inoculation avec Glomus mosseae sur la croissance d'Acacia holosericea en pépinière et après transplantation sur le terrain. Pages 287-293 in Les mycorrhizes, partie intégrante de la plante: biolo-gie et perspective d'utilisation (S. Gianninazzi and Gianninazzi-Pearson, M., eds.). France: Institut national de la recherche agronomique. CTFT (Centre technique forestier tropical). 1988. Faidherbia albida. (Del.) A. Chev. (Synonyme: Acacia albida Del.) Monographie. C.T.F.T., Nogent-sur-Marne. France: CTFT. 72 pp. Dreyfus, B.L, and Dommergues, Y.R. 1981. Nodulation of Acacia species by fast and slow growing tropical strains of rhizobium. Applied Environmental Microbiology 41 (l):97-99. Ducousso, M. 1990. Importance des symbioses racinaires pour l'utilisation des acacias en Afrique de l'Ouest. Thesis, University of Lyon. Lyon, France. 260 pp. Gupta, R.K., Saxena, S.K., and Dutta, B.K. 1973. Germination, seedling behaviour and photomass of some acacias in the nursery stage. Indian Forester 99(6):352-358. Jacques, D. 1986. Rapport sur les recherches effectuées entre le 14 octobre 1984 et le 2 décembre 1985 sur les symbiotes des légumineuses auprès du Centre national de recherches forestieres, Institut sénégalais des recherches agricoles. Nogent-sur-Marne, France: Centre technique forestier tropical. 11 pp. (Limited distribution.) Phillips, J.M., and Hayman, D.S. 1970. Improved procedure for cleaning roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 55:158.
1 Laboratoire de microbiologie forestiere, Direction des recherches sur les productions forestieres (DRPF), Institut senegalais de recherches agricoles (ISRA). Dakar, Senegal. 2 Laboratoire de biologie et physiologie vegetative. ORSTOM. Dakar. Senegal. Ducousso, M., and Colonna, J.P. 1992. Endomycorrhiza infection in young Faidherbia albida: influence on growth and development. Pages 151 -156 I'M Fadherbia albida in the West African semi-arid tropics: proceedings of a workshop, 22-26 Apr 1991, Niamey, Niger (Vandenbeldt. R.J., ed.). Patancheru. A.P. 502 324. India: International Crops Research Institute for the Semi-Arid Tropics: and Nairobi. Kenya: International Centre for Research in Agroforestry. |