![]()
An e-publication by the World Agroforestry Centre
METEOROLOGY AND AGROFORESTRY

|
|
An e-publication by the World Agroforestry Centre |
|
METEOROLOGY AND AGROFORESTRY |
|
|
section 5 : results of agroforestry experiments Water-balance and soil water relations studies in a mixed tree/grass/bare-soil system J.E. Jackson and P J.C. Hamer
Institute of Horticultural Research B. Jackson Sittingbourne College, Sittingboume, Kent, UK Abstract Water balance studies in a mature apple orchard showed that where the grass between the tree rows had been killed by herbicide treatment the efficiency of rainfall in replenishing soil moisture was greatly reduced. The rate of infiltration of water into overall-herbicide treated soil was found to be much less than into soil of grassed alleys. Other experiments have shown soil from grass-free orchard plots to have higher bulk density and lower aggregate stability than that from grassed orchads. Bare soil in the alley-ways is accompanied by lower organic matter content and greatly reduced earthworm activity. It seems likely that the basic cause of the soil structural problems arises from the low rooting density of the trees and their relatively small contribution of organic matter to the soil. The importance of the contribution of the ground cover crop to the maintenance of soil structure in tree plantations and in agroforestry systems in general is dicussed.
Light, water and nutrients can be considered as three "resource pools" exploited by the components of agroforestry systems. The amount of light available at any one site is fixed, so the trees and ground-cover crops share a fixed resource. This is not the case for nutrients where nitrogen-fixing symbionts associated with either the tree or the crop and mycorrhizal infection of the roots can lead to substantial increases in the amounts of available N and P. With respect to water it has long been accepted that the components of a tree/crop mixture can influence total water availability. The emphasis has been on the effects of trees on the pattern of rainfall interception (through the effect of stemflow in, effectively, redistributing rainfall in the horizontal dimension), on evaporative losses (especially from wet leaves), and soil stability. The present paper presents data on the effect of a ground cover crop in a mixed tree and crop system on water availability through its effects in reducing run-off, and discusses this in relation to the relative effects of trees and herbaceous ground cover on the factors controlling soil structure and infiltration of water.
Experimental details Soil water use was measured for the years 1977-83 in a trial to examine the effects of spacing and soil management on the growth and cropping of Golden Delicious apple trees on M.9 rootstocks which had been planted at East Mailing in the winter of 1969/70. The soil is a sandy loam overlying ragstone at 90-120 cm depth and the land slopes from south to north at an angle of 2° to 3°. The trees were in N-S hedgerows (between-row spacings 4.25,3.35 and 2.9 m, within-row spacings 2.7,2.25,1.35 and 0.9 m in all combinations). A belt of soil under the trees was kept clear of weeds by the use of herbicide. This 'herbicide strip' was originally 1 m wide but was gradually increased in width as the trees thickened and by 1983 was approximately 2 m wide. The alley-ways between the herbicide strips were grassed and were mown regularly. In 1977 the soil management regime on two of the four replicates was changed, with the grass in the alleys being eliminated with herbicide and a bare-soil regime, referred to subsequently as overall-herbicide (OH), maintained thereafter. In the treatments reported here, soil water content was measured by neutron probe in the tree spacings of 1.8 m within the tree row and 4.25 m between the rows. Access tubes were arranged in a 50-cm grid from the tree centre, with access tubes parallel to the tree row and 50,100 and 150 cm from it. All access tubes (16 swg aluminium) were installed to bedrock depth. Soil water content of the soil profile was determined using the method of Bell (1973) and the soil moisture deficit was taken as the difference from a reference value when the soil was close to field capacity. The moisture contents for each spacing and soil management type were estimated from measurements for each access tube multiplied by an area factor which represented the area associated with each access tube. Meteorological data were acquired from the records of a standard meteorological site 200 to 250 m from the position of the access tubes. Potential evaporation (Ep) was calculated as for a well-watered grass surface (Penman 1962). The estimated evaporation rates for the orchard plots were calculated from the change in stored soil moisture and precipitation (cf. Hillel 1980). The assumption in this method is that capillary flow and runoff are small compared to evaporation/transpiration. The analyses were restricted to the summer months when, in general, soil moisture deficits were increasing and it could be assumed that drainage out of the root zone was minimal.
Results of the experiment as a whole will be published elsewhere. Only those relevant to the effects of soil management (grass in the alleys versus overall herbicide) are reported here. Initial attempts to relate measured changes in soil moisture to rainfall, potential evaporation and characteristics of the tree and grass canopies showed that these factors did not explain the changes in soil water. The following results illustrate the nature of the discrepancies and the reason.
Table 1 shows changes in soil moisture deficit (SMD) over the period 17 September 1982 to 6 October 1982 during which there was 98 mm of rain (42% of which fell during rainstorms with more than 6 mm per hour precipitation) and 18 mm of potential evaporation; i.e., there was an 'expected' change in SMD of about 80 mm.
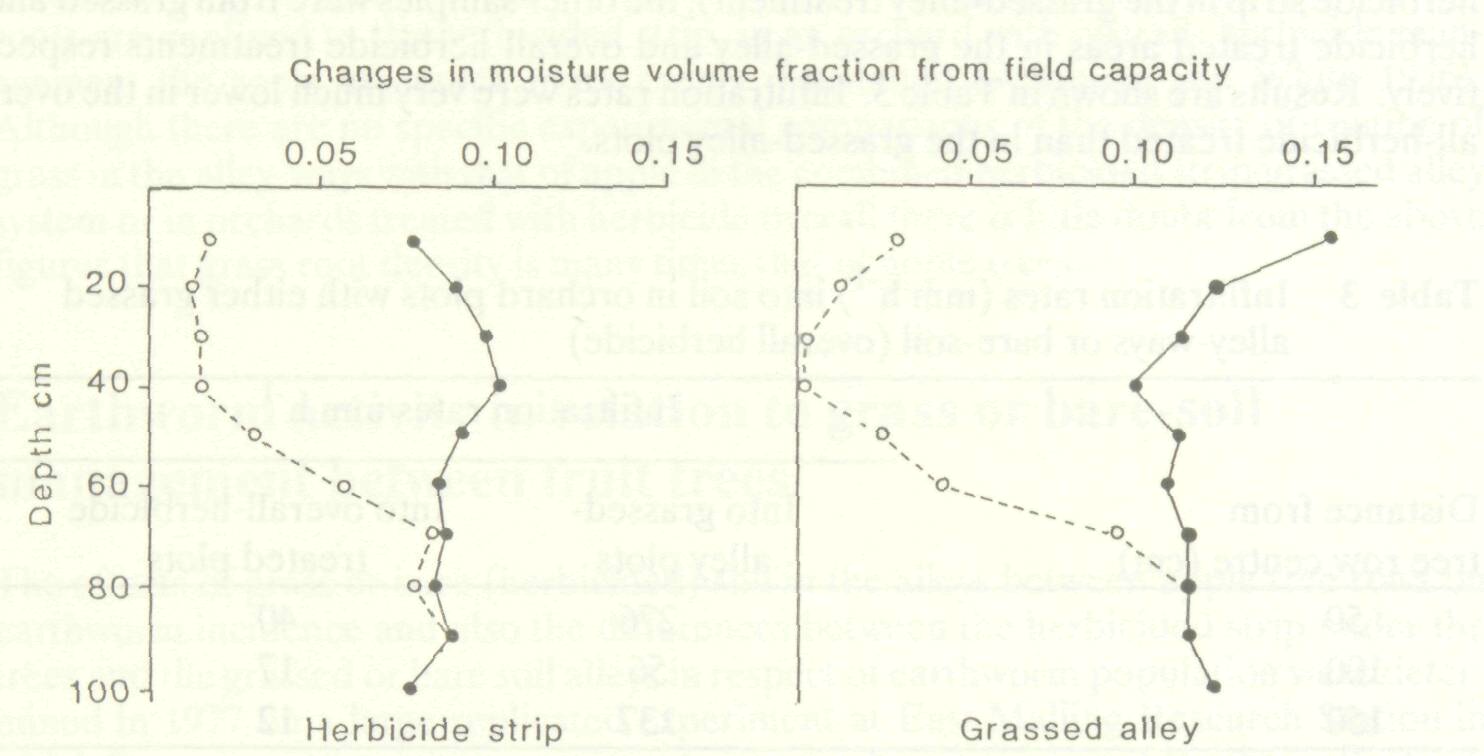
Figure 1 shows the changes in soil moisture deficit with depth between 17 September and 6 October at a distance of 150 cm from the tree trunk. The lack of change at 80 cm and below indicates that there was no drainage to this depth and, by implication, no drainage loss. The decrease of 45 mm in soil moisture deficit instead of the expected 80 mm decrease suggests loss of rain by runoff from the "bare-soil" plots. Similar discrepancies between rainfall and changes in soil moisture deficit under overall herbicide conditions were generally noted and surface runoff was regularly observed. The overall magnitude of the effect was established by looking at changes in soil moisture deficit over 108 separate periods of a week or more over the seven years 1977-1983 (Table 2). These changes were considered as the dependent variable in multiple regression analysis in relation to potential evaporation (Ep) and rainfall (R) according to the equation SMD = a0 + a1 Ep + a2 R (1)
Table 2 Regression analysis of change in soil moisture deficit on potential evaporation (Ep) and rainfall (R) in mm per day according to the equation SMD = a0 + a1 Ep + a2 R for grassed and herbicide treated alley-ways between tree rows 4.25 m apart with trees 1.8 m apart in the rows.
Samples were taken in March 1985 with an Eijkelkamp double ring infiltrometer at distances of 50,100 and 150 cm from the centre of the tree row. The sample at 50 cm from the row centre was, in each case, taken from herbicide treated land (i.e. from the herbicide strip in the grassed-alley treatment), the other samples were from grassed and herbicide treated areas in the grassed-alley and overall herbicide treatments respectively. Results are shown in Table 3. Infiltration rates were very much lower in the overall-herbicide treated than in the grassed-alley plots.
Atkinson and White (1976) compared soil bulk density in the grassed alley-ways of mature (9-year-old) apple orchards with that in corresponding alleys which had been kept free of grass and weeds for the last 3 years by use of herbicides. Soil bulk density was appreciably higher in the bare-soil alley-ways (1.54 g ml-1 as compared with 1.34 in the grassed alleys). Similar results were obtained in other experiments (Atkinson and Herbert 1979). Soil compaction, expressed as 'California bearing ratio' was also much more extensive and severe in orchard alley-ways treated with herbicide than in those which were grassed (Atkinson, Swain and Fricker 1979). O'Kennedy and Robinson (1984) showed that the percentage of water-stable aggregates was much higher in grassed orchard plots (45%) than overall-herbicide treated plots (24%) but that mulching the overall-herbicide treated plots with straw from the time of imposition of the herbicide treatment had some beneficial effect, raising the percentage of water-soluble aggregates to 34.
The most important factors controlling the creation of structural pores in undisturbed soils are plant roots and the larger members of the soil fauna such as earthworms, beetle larvae, ants and termites (Russell 1973). Structural pores, once created, are only stable against wetting if there is some agent holding the soil particles apart against the surface tension forces set up on wetting; among the agencies which maintain stability are the clay fraction, humus and decomposable organic matter. Apple root systems tend to be sparse, reported values of root density expressed as cm root per cm2 soil surface area (LA) being within the range 0.8-23.8 (Atkinson 1980). In contrast the reported values for Graminae are in the range 100 to 4000 and for herbs 52 to 310 (Newman 1969). If rooting density is expressed as cm per cm of soil Lv) the highest value cited for apple by Atkinson (1980) was 0.20, in contrast with the Lvfor grasses of 50 given by Bowen (1984). In an orchard with grassed alleys most of the tree roots are confined to the herbicided strip, in an orchard with overall- herbicide management the roots are much more evenly distributed (Atkinson and White 1976). Although there are no specific experimental comparisons of the density of rooting of grass in the alley-ways with that of apple in the combined herbicided strip/grassed alley system or in orchards treated with herbicide overall there is little doubt from the above figures that grass root density is many times that of apple trees.
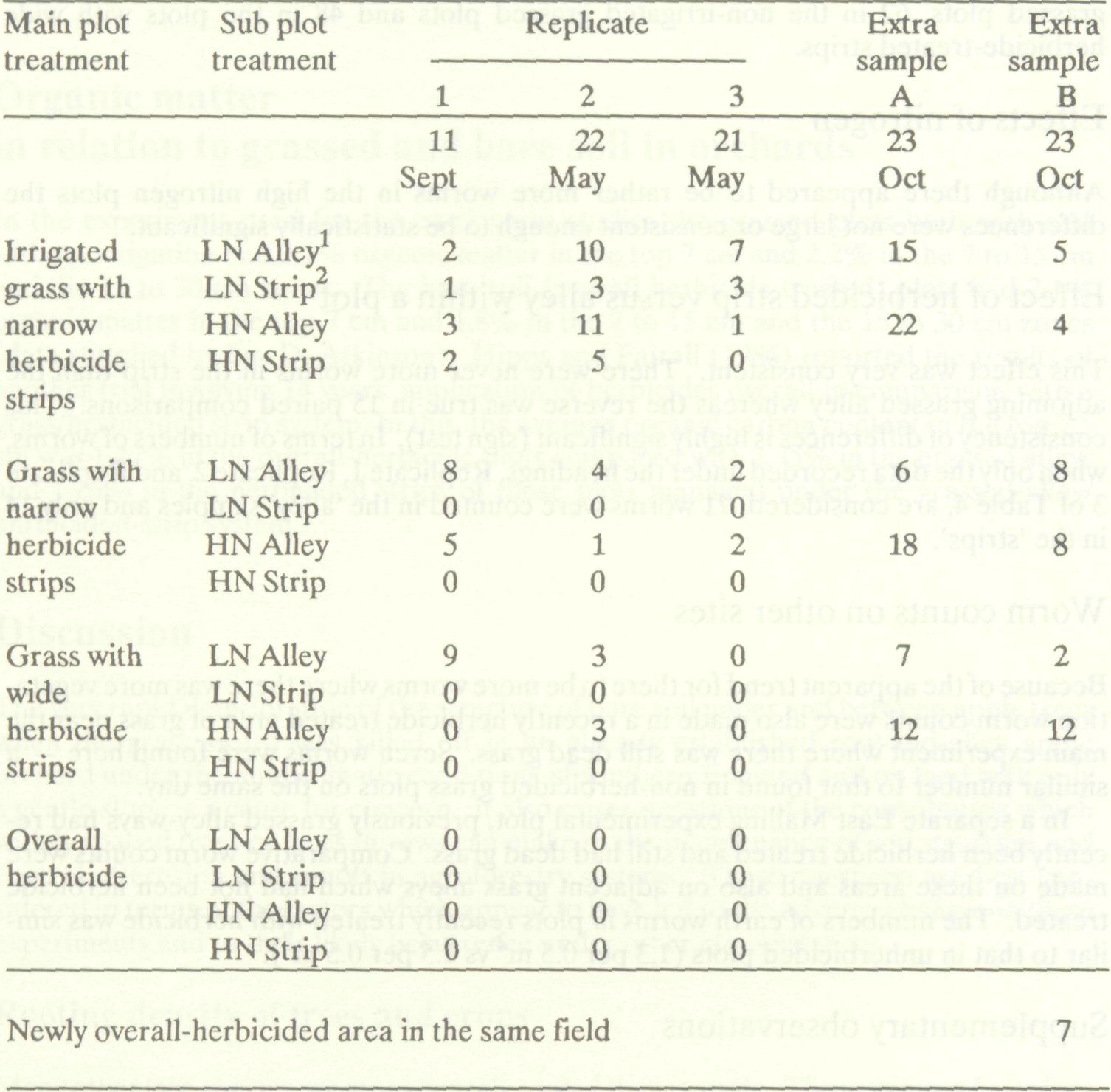
The effects of grass or bare (herbicided) soil in the alleys between apple tree rows on earthworm incidence and also the differences between the herbicided strip under the trees and the grassed or bare soil alleys in respect of earthworm population were determined in 1977 on a large replicated experiment at East Mailing Research Station in which four contrasting management systems were being compared for their effects on tree growth and cropping, when nitrogen fertilizer was applied at either a high or low rate.
The experiment consisted of Cox's Orange Pippin and Spartan apple trees planted in rows 4.5 m apart in December 1972 with the following soil management treatments:
Each of these four 'main plot treatments' was replicated three times and each was subdivided, half being given a high level of nitrogen fertilizer ( potassium nitrate, 189 kg/ha) and half a low level of nitrogen fertilizer (63 kg/ha). This was intended to test whether addition of extra nitrogen fertilizer could overcome the adverse effect of competition by grass for mineral nutrients. Each 'sub-plot' consisted of nine experimental trees of Cox on MM.106 rootstock, spaced 4.5 m apart and surrounded by guard trees, alternately Cox and Spartan at the same spacing. The individual plot size for soil management purposes was therefore 18 m x 18 m. These treatments had all been applied from the time of planting. To test whether some effects of the herbicide treatments on earthworm numbers noted in this study were due to direct effects of the herbicide, or to effects on food supply, earthworm counts were also made on other plots in which the herbicide treatments had been carried out so recently that there was still plenty of dead grass on the soil surface. Earthworms and other soil animals were brought to the surface by the application, using a watering can, of 5 litres of a solution of detergent containing 25 cm3 of a proprietary liquid detergent to the area covered by a 0.5 m2 quadrat. Worm counts were made on all plots in the strip (i.e., within tree rows) and alley (between tree rows) positions. In the irrigated grass and wide strip plots these 'strip' and 'alley' positions corresponded to herbicide-treated and grassed areas respectively. In the plots treated with herbicide overall, both the strip and alley positions were in bare, herbicide- treated soil.
Results are given in Table 4 together with results of ancillary observations. In the overall-herbicide treated plots 'Alley' and 'Strip' all had the same treatment (herbicide) so reflect position, i.e., within-row or between-row, respectively. Table 4 Effects of treatments on number of earthworms per 0.5 m2 soil.
Key LN = Low Nitrogen
Effects of overall herbicide and irrigation treatments Worms were never observed in the overall-herbicide treated plots, although twelve separate assessments were made, and in this consistent absence of worms, these plots were unique. Worms were found in fifteen of the sixteen areas examined on irrigated plots, ten of the sixteen in the grassed plots and seven out of sixteen in the wide herbicided strip plots. Chi-square analysis showed that a significantly higher proportion of the irrigated plots had worms than either the grassed ( P < 0.05 ) or wide herbicide strip plots (P < 0.01). The total number of worms in the different treatments was 94 in the irrigated grassed plots, 62 in the non-irrigated grassed plots and 48 in the plots with wide herbicide-treated strips.
Although there appeared to be rather more worms in the high nitrogen plots the differences were not large or consistent enough to be statistically significant.
This effect was very consistent. There were never more worms in the strip than the adjoining grassed alley whereas the reverse was true in 15 paired comparisons. This consistency of differences is highly significant (sign test). In terms of numbers of worms, when only the data recorded under the headings, Replicate 1, Replicate 2, and Replicate 3 of Table 4, are considered, 71 worms were counted in the 'alley' samples and only 14 in the 'strips'.
Because of the apparent trend for there to be more worms where there was more vegetation worm counts were also made in a recently herbicide treated area of grass near the main experiment where there was still dead grass. Seven worms were found here a similar number to that found in non-herbicided grass plots on the same day. In a separate East Mailing experimental plot, previously grassed alley-ways had recently been herbicide treated and still had dead grass. Comparative worm counts were made on these areas and also on adjacent grass alleys which had not been herbicide treated. The numbers of earth worms in plots recently treated with herbicide was similar to that in unherbicided plots (1.3 per 0.5 m2 vs 1.5 per 0.5 m2 ).
In the experiment used for the earthworm studies the grassed plots both with and without irrigation had 3.1% organic matter in the top 7 cm and 2.2% in the 7 to 15 cm and the 15 to 30 cm zones. The bare soil (overall herbicide-treated) plots had 2.1% organic matter in the top 7 cm and 1.8% in the 7 to 15 cm and the 15 to 30 cm zones (data supplied by Dr. D. Atkinson). Hipps and Fairall (1986) reported the results of another trial involving 14 years' application of herbicides overall in comparison with a grass/herbicided strip system. In this, the average organic carbon content in the top 7.5 cm was 1.47% in the overall-herbicide plots compared with 2.75% in the grassed alleys and 1.26% in the herbicided strips of those plots managed under the grassed-alley/ herbicided-strip system.
The very rapid deterioration of the structure of bare soil under and between apple trees when the grass sward was killed off in an already established row-and-alley apple orchard under the mild climatic conditions of southern England and on land with only a gentle slope is a cause for concern. It also raises questions of the contribution which widely spaced trees or trees in rows can make to the maintenance of soil structure and control of erosion and runoff in agroforestry systems. These questions are best considered in terms of the factors which appear to have led to the adverse changes in these experiments and to their likely occurrence under other circumstances.
Many other tree species are more densely rooted than is apple. The maximum Lv values quoted by Atkinson (1980) are around 0.1 cm root per cm of soil for apple compared with 0.5-0.7 for a range of conifers. Bowen (1984) cites values of from 0.13 to 5.26 in the upper soil zones (0-10 cm and 0-15 cm respectively) for conifers and up to 8 in the 0-10 cm zone for Eucalyptus. It would seem from this that the root system of other trees might make a much greater contribution to the creation of structural pores than do apple root systems. However, even the highest Lv values which Bowen gives for trees are many times lower than the value of 50 in the 0-15 cm zone which he gives for grasses and at the lower end of the range of 5 to 25 which he gives for cereals. Newman (1969) gives the range of LA values reported for Graminae as 100-4000, other herbs 53-310 and woody plants 5-110. The contribution of tree roots to creating good soil structure therefore seems likely to be small relative to that of grasses and crop plants. The inadequacies of trees in this respect will be accentuated in the early years of tree plantations unless there is a ground-cover crop.
The results in the experiments at East Mailing are consistent with the incidence of earthworms being dependent on the availability of food supplies. Their numbers were approximately proportional to the vigour of grass growth, with most being found from under the irrigated grassed plots, next most from the unirrigated grass and none from the overall-herbicided plots. It is unlikely that any direct effect of the herbicides in poisoning earthworms was involved. Worms and worm casts were frequent in newly herbicide-treated land with dead grass on the surface. Results from Rothamsted, England, showed that killing pasture with herbicide and slit-seeding a cereal crop without further cultivation gave a much higher population of earthworms in the first year out of pasture than did traditional ploughing (Russell 1973). The absence or greatly reduced numbers of earthworms in the orchard herbicided strips, which were 1.7 m or less in width, indicates the specificity of the link between above-ground vegetation and earthworm activity below ground, presumably as a result not only of leaf litter but also of the turnover of roots. It seems reasonable to suppose that a similar link will apply in all temperate and tropical areas where earthworms are an important component of the soil ecology. An obvious deduction is that any portion of the soil surface kept bare of vegetation in agroforestry systems or the early stages of forestry or tree crop plantations will be accompanied by absence of earthworms and potential development of poor soil structure and reduced water infiltration.
The trees in apple orchards, which usually have LAIs of less than 2.5 (Jackson 1980) and have the sparse root systems discussed previously can be expected to return less organic matter to the soil than many others. Broadleaf forest trees have LAIs of 6 to 7 (Kira and Kumura 1983), rubber an LAI at canopy closure of 6.3 (Moraes 1977); and cacao LAIs varying from 3.7 to 5.7 (Alvim 1977). On the assumption that the root production and turnover in the soil is proportional to these LAIs (which may not be valid), the organic matter inputs into the soil from forest trees and crops such as cacao and rubber at maturity should be higher than those from apple. They will still, however, be low compared with those from grasses or grain crops (leaving root systems in the soil) with higher LAIs and net production efficiencies (Table 2; Kira and Kumura 1983). This will be particularly so in the early years of tree establishment when net dry matter productivity per hectare can be very low. There are conflicting data on soil organic matter changes under different tree crops but in some cases at least, even under good management topsoil organic carbon has been reported to fall following the establishment of plantation tree crops, e.g., by 41% in 14 years in an oil palm plantation (Sanchez et al. 1985). A major factor controlling the organic matter inputs by trees and ground-cover plants into the soil is the actual destination of the organic matter produced. Apple orchards subjected to herbicide management suffer because the leaves shed on to the soil tend to blow off and accumulate against boundary fences. Litter removal from teak plantations may be the main reason for runoff and erosion where this occurs (Lundgren 1978). Apparently high efficiency in the use of the different products of trees for fodder, fuel and timber, may result in them making relatively little contribution to the maintenance of soil organic matter status.
At least under some conditions there is marked loss of soil structure leading to considerable reduction in the infiltration of rain water, and severe loss of this by runoff, in mature apple orchards in which the grass cover-crop in the alleys has been removed by herbicide use. This loss of structure is accompanied by, and probably caused by, reductions in the total root occupancy of the soil, in organic matter content and in earthworm activity as compared with grassed orchard soils. These results are probably relevant to row-and-alley and other agroforestry systems in general. They may be relatively extreme because of the generally low Lv values of apple root systems and the low LAIs, hence potential biomass production, of apple orchards. The problems did, however, develop within a short space of years on land with only a gentle slope in the mild climate of southern England. This suggests that serious attention should be given to the ground- cover crops within all agroforestry systems, in which the trees, by definition, do not provide a dense closed canopy, if problems of soil structure, runoff and erosion are to be avoided. It is of interest that Gary and Evans (1972) in an experiment on the effect of soil management regimes under citrus trees found the highest infiltration rates to be into permanent sod, followed in turn by winter cloves, winter tick-bean and bare surface chemical weed control treatments.
Alvim, P. de T. 1977. Cacao. In P. de T. Alvim and T.T. Kozlowski (eds.), Eco physiology of tropical crops. New York, San Francisco, London: Academic Press. Atkinson, D. 1980. The distribution and effectiveness of the roots of tree crops. In J. Janick (ed.), Hortic. Rev.. Vol. 2. Atkinson, D. and R.F. Herbert. 1979. A review of long term effects of herbicides: effects on the soil with particular reference to orchard crops. Ann. Appl. Biol. 91:125-129. Atkinson, D., R.W. Swain and D. Flicker. 1979. The effect on soil condition of herbicide use in fruit plantations. In A. Banin and U. Kafkafi (eds.), Agrochemicals in soils. Berlin: Springer Verlag. Atkinson, D. and G.C. White. 1976. Soil management with herbicides the response of soils and plants. In Proceedings 1976 British Crop Protection Conference -Weeds. 3: 873-884. Bell, J.P. 1973. Neutron probe practice. Report No. 19, Institute of Hydrology. Bowen, G.D. 1984. Tree roots and the use of soil nutrients. In G.D. Bowen and E.K.S. Nambiar (eds), Nutrition of plantation forests. London: Academic Press. Cary, P.R. and G.N. Evans. 1972. Long term effects of soil management treatments on soil physical conditions in a factorial citrus experiment. J. Hort. Sci 47:81-91. Hillel, D. 1980. Applications of soil physics. New York: Academic Press. Hipps, N.A. and G.B.N. Fairall. 1986. Effects of soil management treatments on soil organic carbon content. Report of the East Mailing Research Station for 1985. p. 159-160. Kira, T. and A. Kumura. 1983. Dry matter production and efficiency in various types of plant canopies. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi, Kenya: ICRAF. Lundgren, B. 1978. Soil conditions and nutrient cycling under natural and plantation forest in Tanzanian highlands. Report on forest ecology and forest soils, No. 31. Uppsala: Swedish University of Agricultural Sciences. Moraes, V.H.F. 1977. Rubber. In P.de T. Alvim and T.T. Kozlowski (eds.), Eco-physiology of tropical crops. New York, San Francisco, London: Academic Press. Newman, E.I. 1969. Resistance to water flow in soil and plant. 1. Soil resistance in relation to amounts of root: theoretical estimates. J. Appl. Ecol. 6:1-12. O'Kennedy, N.D. and D.W. Robinson. 1984. Further results from trials with overall herbicides in apples at Eallygagin. Aspects Appl. Biol., Weed control in fruit crops. 8:169-177. A.A.B., N.V.R.S. Wellesbourne. Penman, H.L. 1962. Woburn irrigation, 1951-59. I. Purpose, design and weather. J.Agric.Sci. 58: 343-348. Russell, E.W. 1973. Soil conditions and plant growth. London: Longman. Sanchez, PA., CA. Palm, C.B. Davey, L.T. Szott and C.E. Russell. 1985. Trees as soil improvers in the humid tropics? In M.G.R. Cannell and J.E. Jackson (eds.), Trees as crop plants. Institute of Terrestrial Ecology, Natural Environment Research Council. |