![]()
An e-publication by the World Agroforestry Centre
METEOROLOGY AND AGROFORESTRY

|
|
An e-publication by the World Agroforestry Centre |
|
METEOROLOGY AND AGROFORESTRY |
|
|
section 2 : basic principles Tree and crop selection and management to optimize overall system productivity, especially light utilization, in agroforestry J.E. Jackson
Institute of Horticultural Research Abstract Optimizing management of agroforestry systems necessitates maximizing the efficiency of use of light resources. This involves recognition of the light requirements of the trees and crops involved and the factors influencing light interception by the trees and penetration to the ground crop. There are four main types of crop light response:
Where shade is required (Type D) then sparsely foliated trees giving uniform shade over most of the ground may be needed and such a shading pattern may also be advantageous with Type B ground crops. In general, however, the most important requirement of the tree crop is that it maximizes the yield of economic product per unit of cast shade. Fruit trees tend to be of Type C and their density and volume of canopy can often be reduced with little or no loss of yield and, by so doing, let more light to the ground crop. This can be achieved either genetically, in particular by selecting for columnar types, or by the pruning techniques used. Forest tree genotypes with a high ratio of timber production to total tree volume, hence cast shadow, are also available and would be particularly useful in agroforestry. Where fuelwood or fodder trees are grown in agroforestry blocks under subsistence farming or smallholder circumstances a key objective must be to sustain production. Harvesting methods which have only a minimal effect on light interception are the key to this. Removing lower branches will have little adverse effect on subsequent light interception and canopy productivity. Reducing canopy dimensions, especially by harvesting outer rows, will have a much greater adverse effect on productivity per unit of fuel or fodder harvested. Removal of individual trees throughout the block will have effects part-way between those of branch removal and those of reducing the dimensions of the wooded area.
Agroforestry systems incorporating a range of tree and crop species offer much more scope for useful management of light interception and distribution than do monoculture forests and agricultural crops. The potential benefit as a result of combining field crops with trees is obvious from consideration of the waste of light resources experienced in orchards and tree-crop plantations. Trees are commonly planted at spacings appropriate to their size at maturity. This inevitably leads to under-use of available light in the early years after planting. Even modern, productive apple orchards can intercept as little as 30% of the available light at five years of age (Jackson 1980). A coconut plantation that intercepted 90 percent of the available light at maturity intercepted 50% when three years old and 70% when five years old (Nelliat et al. 1974; reported by Nair 1983). Even at maturity many tree crops transmit a high proportion of the available light because experience has shown that it is not beneficial to have the trees denser or more closely spaced. Apple orchards seldom intercept more than 70% of the available light, those which do so being too dense and shaded for profitable fruit production (Jackson 1980). Twenty-year-old tall coconut palms in the Russell Islands were found to transmit 40-70% of the available light, depending on planting density (Smith and Whiteman 1983) and a dense dwarf palm plantation transmitted 20%. Okigbo and Greenland (1976) and Okigbo (1980) identified more efficient use of light resources by plants of different heights and canopy structures as one of the advantages to be gained by growing crops in mixed stands. The way in which this is achieved in one traditional agroforestry system, the village-forest-garden system of Indonesia, was described by Michon (1983). In these forest-gardens, the cultivated crop species, from small herbs to taller emergent trees, form a layered structure giving a gradient in light and humidity exploited by plants according to their ecological requirements. It is reasonable to suppose that successful systems of tropical agroforestry evolved along the lines described by Bunting (1980) for tropical mixed cropping in general. As soon as a gap appears in the mixture the farmer, or his wife, puts something in it. A body of knowledge is thus built up as to the combinations of available plants which most nearly meet the farmer's requirements. The effective application of the much more rapid and powerful modern scientific methods of improving agricultural productivity to these mixed crop systems depends on understanding the crop interactions, particularly in relation to use of light, water and nutrient resources and developing appropriate management strategies. This paper concentrates on light utilization in agroforestry systems and the ways in which this can be optimized.
The characteristics feature of light interception and penetration in discontinuous canopies such as those in agroforestry is that it cannot be calculated as a simple function of vertically-summed leaf area index (L) and a light extinction coefficient (K) by the usual equation: I / Io = e-KL (1) where I is light intensity beneath an LAI of L and I0 is light intensity above the canopy. Instead the transmission through the (discontinuous) tree canopy must include a component representing light which misses the trees altogether by penetrating directly to the ground crop through gaps between the trees or rows of trees. If such transmission is designated Tf then the total light transmitted to the ground or field crop surface (T) expressed as a decimal fraction of light above the trees is as shown in Equation (2) (Jackson and Palmer 1979; Jackson 1983): T=Tf + (1-Tf)e-KL (2) In this equation Tf represents the fraction of the ground surface which would still receive light if the trees were 'solid' or non-transmitting to light and the second term represents light actually transmitted to the ground through the tree canopy with L' being L/(l-Tf ). The corresponding equation for fractional interception by the trees ( F ) is: F=Fmax-Fmax e-KL (3) where Fmax is the decimal fraction of the available light which would be intercepted if the trees were 'solid'. From these equations it can be seen that:
Using a modified Norman and Welles (1983) radiation interception model they calculated that under a particular set of conditions for Pinus radiata in New Zealand increasing the crown height to give a 77% increase in crown volume, with a correspondingly reduced foliage density, increased photosynthate production per tree and per unit ground area by 12%.
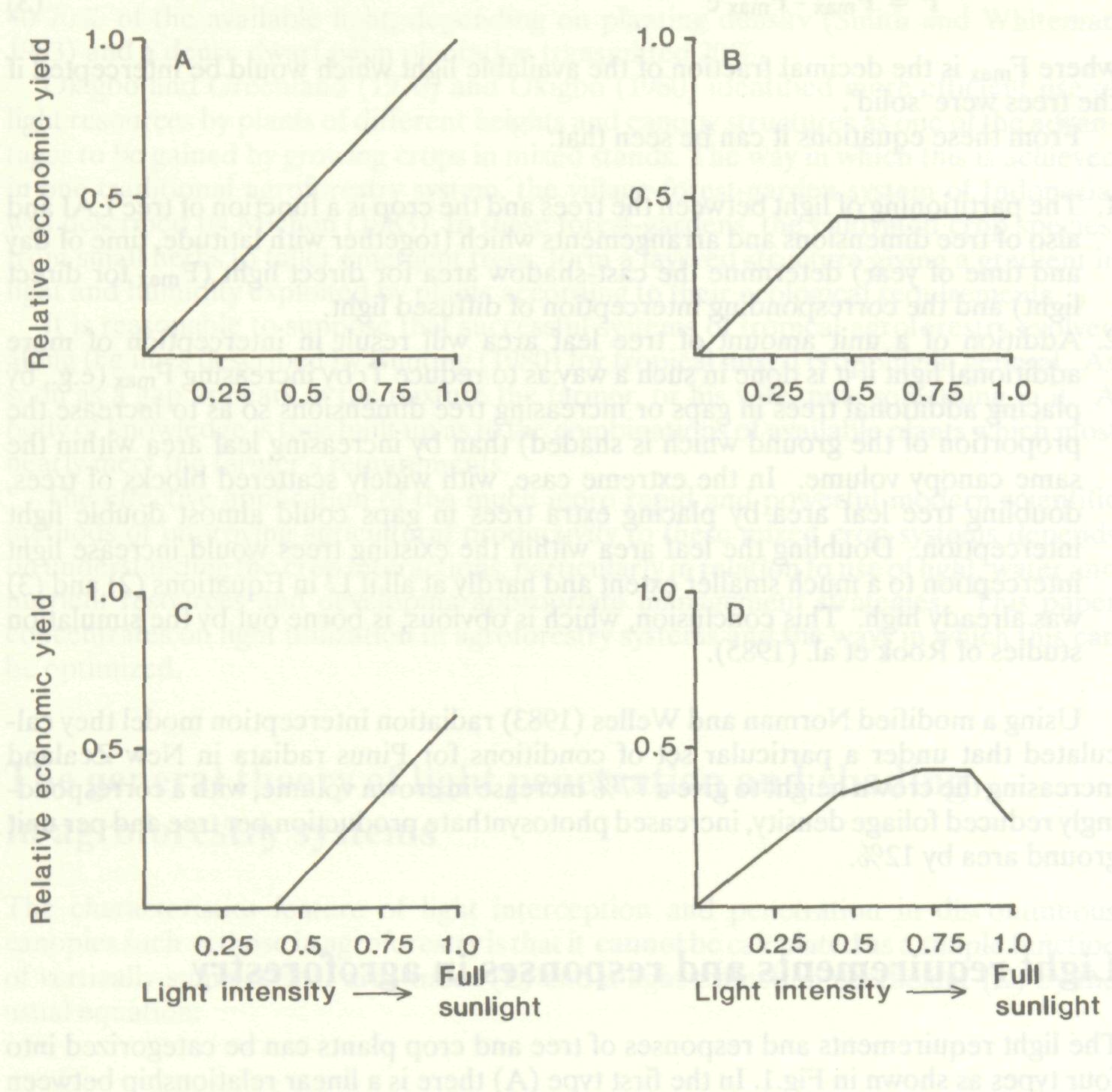
The light requirements and responses of tree and crop plants can be categorized into four types as shown in Fig.1. In the first type (A) there is a linear relationship between intercepted light or light intensity with the regression line going through the origin. There are many examples of dry matter production by crop plants being proportional to intercepted radiation, particularly early in the season (Biscoe and Gallagher 1977; Monteith 1977a). This does not necessarily imply a linear relationship between canopy productivity and light intensity because at least some of the variation in radiant energy interception in the data used was a consequence of variations in percentage effective ground cover (percentage of available light intercepted) rather than in light intensity. There is, however, also direct evidence that net canopy photosynthesis can be as linearly dependent on irradiance changes during a day (Biscoe and Gallagher 1977).
Monteith (1972) cites crops such as maize, bulrush millet and tropical grasses in general as having this type of non-saturated light response. Subsequently Monteith (1977b) attributed this to C4 plants in general. The second characteristic type of crop x light relationship (B) is that in which crop canopies show light saturation at less than full sunlight intensity. In this case net photosynthesis or crop growth increases with light intensity up to a certain level and then stays at this plateau rate irrespective of further increases in irradiation (Biscoe and Gallagher 1977; Sale 1974). Mott and Popenoe (1977) show this response curve as typical of tropical legumes and Monteith (1972) shows a rather less flattened one for tropical species such as cassava and beans. The third type of relationship (C) is one in which there is little or no economic yield at below a threshold level of light intensity although yield can be a linear function of light intensity above this point. Cannell (1983) suggested that fruit and seed-yielding crops in general are intolerant of shade. It seems that the processes involved in flower and fruit bud formation, fruit set, fruit growth and the development of economically important visual attributes such as red skin pigmentation (anthocyanin formation) require levels of insolation much higher than those needed for vegetative growth. This was shown by within-canopy studies and artificial shading experiments on apple in which progressive increases in shade which had little effect on vegetative growth had dramatic effects on fruit yield and quality (Jackson and Palmer 1977a, 1977b; Jackson et al. 1977). From this and other work it was concluded that canopy zones receiving less than 50% of full daylight were effectively incapable of producing good quality apples and studies on peach planting systems suggest that fruit formation in this stone-fruit crop is at least as shade intolerant. The final category (D) includes trees and crops which actually benefit from shading. Huxley (1967) showed by direct experimentation that Robusta and Arabica coffee seedlings increased in dry weight more rapidly when grown under 27, 38 or 54% full daylight than when grown in the open. This phenomenon appears widespread among tree seedlings. A rather more surprising result, in view of the generally adverse effects of shade on fruiting, was the increase in yield of top quality fruits of peppers in Israel which Rylski (1986) achieved by artificial shading. When imposed in summer, low levels of shade (12 and 26%) gave yield improvements of more than 62% over the unshaded controls while reducing light by 47% gave 30% more yield than control.
Selection and management of 'high light requirement' fruit tree crops There is ample evidence that seed and fruit producing trees are often of Type C as defined above. They cannot produce their crop in the more shaded parts of the trees and consequently there has been a pronounced move towards using very dwarfed trees giving shallow, well-illuminated cropping canopies in the commodities which have had the greatest research input. The change in apple production has been particularly dramatic with large trees replaced by more productive shallow cropping canopies of little more than a metre in depth and with orchard LAIs of around 2 or less (Jackson 1980; Jackson and Middleton 1987) compared with old style large tree orchards with LAIs of 5 or more. The productivity gain in the sole crop orchard is a result of better illumination of the fruiting wood, easier and cheaper management as a result of the reduced tree size (Jackson 1985) and a lower respiratory load in relation to photo-synthetic productivity. In the agroforestry situation there would be a tremendous additional advantage of eliminating the non-productive zones of fruit- and seed-producing trees in that this would enable light currently intercepted unprofitably by the lower and inner parts of such trees to be available for growing understorey crops. The simplest approach to meeting the specific agroforestry objective is to produce vertical columnar trees capable of producing fruits all the way down to ground level and so contained in their lateral growth that they bear fruits right back to the tree trunk, i.e. there is no unproductive inner zone. Such trees would minimize Fmax for any given level of fruit production, minimize light interception per unit of tree LAI, and maximize availability to the inter-crop at ground level. There are two ways of achieving such tree form, by selection and breeding; and by pruning.
Variation in branching habit and tree form is commonplace within both coniferous and deciduous tree species, arising by mutation, but it is only recently that this variation has been exploited in a systematic way other than for purely ornamental purposes. Two common mutant forms, those generally referred to as 'pendula' and 'columnaris' can have the narrow-crowned habit which minimizes their cast shadow areas. 'Spur-types' with the main branches clothed in fruiting spurs rather than long lateral branches are also commonly found, e.g., in apple and in peach, an outstanding example being Red Delicious apple of which spur-types predominate. Such trees are, by and large, of conventional form even though the ratio of fruit yield to tree cast shadow area is obviously greatly increased. A much greater potential for radical change in tree form has come with the identification of the gene for compact, columnar, non- branching habit and its use in breeding programmes (Tobutt 1985). Trees of this type are essentially upright poles covered with fruiting spurs without any lateral vegetative branches. The first varieties from these programmes are just now becoming commercially available. A rather similar habit is available within Prunus, although so far only exploited in the ornamental cherry Amanagawa. There are also prospects of compact types of a number of other fruit species (Alston and Spiegel-Roy 1985).
Depending on natural tree branching and fruiting patterns two methods suggest themselves. Where the fruit trees crop on young wood, e.g., one-, two- and three-year-old wood, they can be pruned to produce a vertical axis with cropping laterals which are removed by cutting back to the trunk as soon as they surpass the desired maximum age. This is the basis of the 'pillar' and the 'slender spindle bush' (Wertheim 1978) and also of the 'axe' which is being used for both apples and peaches in France (Lespinasse and Delort 1986). The major weakness exhibited by 'pillar' systems in the United Kingdom and 'axe' systems in France is that, where the trees are planted relatively close together in hedgerows only a few metres apart and growth is vigorous, the lower parts of the trees become excessively shaded by the adjacent hedgerows, are unable to produce good quality fruits and may even fail to regenerate lateral branches after pruning back to the trunk. Control of the vigour of growth of the top of the trees by careful pruning is essential to prevent this happening if vigour is not naturally controlled by genotype (scion and rootstock) or soil. In the agroforestry situation this problem is much less likely to occur because the trees or hedgerows can be spaced so widely apart that even the lowest parts of the tree receive ample sunlight for production of good quality fruits and for branch regeneration. Where the fruit trees crop best on older spurs then either:
Selection and management of other tree crops in agroforestry situations Some forest trees as well as fruit trees are shade-intolerant (Perry 1985) and Assmann (1970) concluded that mixing shade-tolerant and shade-intolerant trees led to increased productivity compared with growing intolerants alone. For example, the yield of Pinus sylvestris - Fagus sylvatica mixtures was up to 24% greater than that of pure P. sylvestris. Shade-intolerant timber trees can be regarded as ideal partners for understorey crops in the same way as fruit trees and palms and the same design requirements are relevant. Even when the trees are Type A in their light response it is clearly advantageous in an interspersed agroforestry situation for the timber trees to have a maximum ratio of economic yield (trunk) to tree volume and cast-shadow area ( Fmax ). Essentially to achieve this we need trees with strong genetically determined apical dominance (or apical control). This tree type has been sought for very different reasons in the Finnish forest tree breeding programme. In this the ideotype for conifers has stems which are straight, have small taper, thin bark, good quality timber and rapid height and diameter growth, and branches which are slow growing, are consequently small in length and diameter and subtend angles at about 90 degrees with the stems (Karki and Tigerstedt 1985). Such trees could be very suitable for row and alley cropping (see below). The Finnish selection and breeding programmes have produced narrow crowned trees with a high, and highly heritable, ratio of standing stemwood to total above-ground fresh weight. Among the species with narrow crown forms are Picea abies, Pinus sylvestris and Betula pendula. It is noteworthy that narrow-crowned clones occurred naturally as did the Wijcik mutant of Mclntosh which is the source material for compact apple tree breeding. There seems every possibility that a systematic search for narrow-crowned or columnar types of relevant tree species for agroforestry would result in the rapid identification of suitable clones. This argument is probably relevant in all agroforestry situations where timber is an important product. It is likely to be most difficult to put into effect where the ground crops is of Type D with a specific shading requirement. In general it is much easier to ensure an even low level of irradiation in the understorey if the trees have high Fmax and low leaf area per unit canopy volume, grown as flat-headed ( umbrella-like ) trees or as very tall thin hedgerows, than if they are dense and columnar with low Fmax and high density of shading although this can be achieved in row and alley systems with appropriate dimensions and row orientations (Jackson and Palmer 1988). The ideal understorey crop or alley crop to combine with dense columnar spaced trees is one with an Type A light response so that it can make use both of very high irradiances in unshaded areas and low irradiances close to the trees.
It is assumed that these have a Type A light response, with some economic product, in the form of branches and leaves, being produced even in shaded situations and trunk thickening and upward growth being a function of total light interception. There is not likely to be any intrinsic advantage from growing such trees in close, intercropping association with crops. Indeed their yield of vegetative dry matter per unit ground area may well be higher if they are grown in solid blocks with a degree of 'density stress' (Cannell 1983). If the trees are grown in such blocks, or at least in multi-rows, their productivity will be influenced by the following:
In general, the techniques of clear-felling of forest trees and even of coppicing are largely determined by the specification of the end product, e.g., in terms of sawnwood potential, pole characteristics, etc., and by the economics of harvesting on a large scale. In the small-scale agroforestry situation it may well be possible to treat fodder and fuel-wood as regular crops from the trees while maintaining them as photosynthetically efficient canopies. In this context it is worth noting that fruit trees, once mature, are capable of putting 70% of their dry matter increment into harvested fruit each year (Jackson 1985) and data presented by Barlow and Smith (1971) shows up to 77.5% of the total annual increment going into harvested fruit and deciduous leaves with only 22.5% going into root and trunk and branch tissues. If such a ratio of consumable to structural product could be obtained in fodder trees by appropriate selection and management then sustained annual harvesting of only that production which is surplus to the trees' maintenance needs could well replace the longer term cycles of production (Cannell 1983) which are currently common.
Although it is possible to think of available radiant energy being partitioned up between the tree crop and the ground crop the proportionate partitioning will only have a corresponding effect on ground crop yields if this crop is Type A in its light responses. In all other cases the pattern of shade distribution over the ground could be of critical importance. Jackson and Palmer (this volume, Section 5) show that row and alley systems can result either in relatively uniform shade across the alley or in a combination of dense shade near the trees and little shade in the alley centre or adjacent to a south-facing row (if north of the equator). Moreover, the shade pattern can change with time of year. The uniformity of shade cast by scattered or evenly distributed trees will depend on the height of their foliage above the ground as well as on its LAI. Under these circumstances the efficiency with which a crop can utilize the light which is not intercepted by the trees will depend on the combination of crop requirements and light intensity pattern over space and over time. In order to assess the magnitude of these effects it will be necessary to obtain data on shade responses under the appropriate environmental conditions. An indication of the likely crop-specificity can be obtained from the work of Willey and Holliday (1971a, 1971b). They showed the yield of barley to be reduced by shading during the ear development phase but not by shading during the subsequent grain-filling period; whereas the yield of wheat was reduced by shading at either of those times. Yoshida and Parao (1976), in experiments involving shading to 25, 50 and 75% of full daylight, found the vegetative growth of rice to be almost unaffected by shade but the reproduction and ripening phases to show a linear dependence of yield on light intensity. Where shade is of positive benefit, as in protecting peppers from sunscald, it is unlikely that a combination of exposure to full sunlight for part of the day coupled with deep shade for another part of the day as is achieved with some row and alley systems will be of equal value to an equivalent average reduction in light as a result of a virtually continuous sparse overhead canopy. A combination of shade trials and computer modelling of light interception and distribution could be very valuable as a supplement and a guide to direct experimentation to optimize agroforestry design and management and the selection of appropriate trees and crops.
Alston, F.H. and P. Spiegel-Roy. 1985. Fruit breeding strategies, achievements and constraints. In M.G.R. Cannell and J.E. Jackson (eds.) Attributes of trees as crop plants. Institute of Terrestrial Ecology, Natural Environment Research Council. Assmann, E. 1970. The principles of forest yield study. Oxford: Pergamon. Bunting, A.H. 1980. The future of research on mixed cropping in tropical agriculture. In R.G. Hurd, P.V. Biscoe and C. Dennis (eds.), Opportunities for increasing crop yields. Boston, London and Melbourne: Pitman. Barlow, H.W.B. and J.G. Smith. 1971. Effect of cropping on growth of the apple tree. Report of the East Mailing Research Station for 1970. p. 52. Biscoe, P. and J.N. Gallagher. 1977. Weather, dry matter production and yield. In J J. Landsberg and C.V. Cutting (eds.), Environmental effects on crop physiology. London, New York and San Francisco: Academic Press. Cannell, M.G.R. 1983. Plant management in agroforestry. In P. A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Huxley, PA. 1967. The effects of artificial shading on some growth characteristics of Arabica and Robusta coffee seedlings. I. The effects of shading on dry weight, leaf area and derived growth data. J. Appl. Ecol. 4: 291-308. Jackson, J.E. 1980. Light interception and utilization by orchard systems. In J. Janick (ed.), Hortic. Rev. Vol. 2. Jackson, J.E. 1983. Light climate and tree-crop mixtures. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Jackson, J.E. 1985. Fruit orchard design. In M.G.R. Cannell and J.E. Jackson (eds.), Attributes of trees as crop plants. Institute of Terrestrial Ecology, Natural Environment Research Council. Jackson, J.E. and S.G. Middleton. 1987. Orchard design for maximum yield and quality. Proceedings of Conference, La coltura del Melo verso gli anni '90'. Societa Orticola Italiana. Jackson, J.E. and J.W. Palmer. 1977a. Effects of shade on the growth and cropping of apple trees. I. Experimental details and effects on vegetative growth. J. Hortic. Sci. 52:245-252. Jackson, J.E. and J.W. Palmer. 1977b. Effects of shade on the growth and cropping of apple trees. II. Effects on components of yield. J. Hortic. Sci. 52:253-266. Jackson, J.E., J.W. Palmer, MA. Perring and R.O. Sharpies. 1977. Effects of shade on the growth and cropping of apple trees. HI. Effects on fruit growth, chemical composition and quality at harvest and after storage. J. Hortic. Sci. 52: 267-282. Jackson, J.E. and J.W. Palmer. 1979. A simple model of light transmission and interception by discontinuous canopies. Ann. Bot. (Lond.) 44: 381-383. Karki, L. and P.M A. Tigerstedt. 1985. Definition and exploitation of forest tree ideotypes in Finland. In M.G.R. Cannell and J.E. Jackson (eds.), Attributes of trees as crop plants. Institute of Terrestrial Ecology. Natural Environment Research Council. Kira, T. and A. Kumura. 1983. Productivity and plant canopies. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Lespinasse, J.M. and J.F. Delort. 1986. Apple tree management in vertical axis: appraisal after ten years of experiments. Acta Hortic- (The Hague) 160:139-155. Michon, G. 1983. Village-forest-gardens in West Java. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Monteith, J.L. 1972. Productivity in tropical ecosystems. J. Appl. Ecol. 9: 747-766. Monteith, J.L. 1977a. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 281: 277- 294. Monteith, J.L.1977b. Climate. In P. de T. Alvim and T.T. Kozlowski (eds.), Ecophysiology of tropical crops. New York, San Francisco, London: Academic Press. Mott, G.O. and H.L. Popenoe. 1977. Grasslands. In P. de T. Alvim and T.T. Kozlowski (eds.), Ecophysiology of tropical crops. New York, San Francisco, London:Academic Press. Nair, P.K.R. 1983. Agroforestry with coconuts and other tropical plantation crops. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Nelliat, E.V., K.VA. Bavappa and P.K.R. Nair. 1974. Multistoreyed cropping - a new dimension of multiple cropping in coconut plantations.World Crops. 26:262- 266. Norman, J.M. and J.M. Welles. 1983. Radiative transfer in an array of canopies. Agron. J. 75: 481-488. Okigbo, B.N. 1980. The importance of mixed stands in tropical agriculture. In R.G. Hurd, P.V. Biscoe and C. Dennis (eds.), Opportunities for increasing crop yields. Boston, London and Melbourne: Pitman. Okigbo, B.N. and D J. Greenland. 1976. Intercropping systems in tropical Africa. In R.I. Papendick, P.A. Sanchez and G.B. Triplett (eds.), Multiple cropping. American Society of Agriculture Publication No.27. Madison: American Society of Agronomy, Crop Science Society of America; Soil Science Society of America. Perry, D.A. 1985. The competition process in forest stands. In M.G.R. Cannell and J.E. Jackson (eds.), Attributes of trees as crop plants. Institute of Terrestrial Ecology, Natural Environmental Research Council. Rook, D.A., J.C. Grace, P.N. Beets, D. Whitehead, D. Santantonio and H.A.I. Madgwick. 1985. Forest canopy design : biological models and management implications. In M.G.R. Cannel and J.E. Jackson (eds.), Attributes of trees as crop plants. Institute of Terrestrial Ecology, Natural Environment Research Council. Rylski, 1.1986. Improvement of pepper fruit quality and timing of harvest by shading under high solar radiation conditions. Acta Hortic. 191: 221-228. Saeki, T. 1960. Inter-relationships between leaf amount, light distribution and total photosynthesis in a plant community. Bot. Mag. Tokyo. 73: 55-63. Sale, P J.M. 1974. Productivity of vegetable crops in a region of high solar input. III. Carbon balance of potato crops. Aust. J. Plant Physiol. 1: 283-296. Smith, MA. and P.C. Whiteman. 1983. Evaluation of tropical grasses in increasing shade under coconut canopies. Exp. Agric. 19:153-161. Tobutt, K.R. 1985. Breeding columnar apples at East Mailing. Acta Hortic. 159: 63-68. Wetheim, S.J. 1978. Pruning of slender spindle type trees. Acta Hortic. 65:173-180. Willey, R.W. and R. Holliday. 1971a. Plant population and shading studies in barley.J.Agric. Sci. Camb. 77: 445-452. Willey, R.W. and R. Holliday. 1971b. Plant population, shading and thinning studies in wheat.J.. Agric. Sci. Camb. 77:453-461. Yoshida, S. and F.T. Parao. 1976. Climatic influence on yield and yield components of lowland rice in the tropics. In Climate and Rice. Los Banos, Laguna, Philippines: Int. Rice Res. Inst. |