![]()
An e-publication by the World Agroforestry Centre
METEOROLOGY AND AGROFORESTRY

|
|
An e-publication by the World Agroforestry Centre |
|
METEOROLOGY AND AGROFORESTRY |
|
|
section 5 : results of agroforestry experiments Microclimatic modification in intercropping and alley-cropping systems* J.E. Corlett and C. K. Ong
International Crops Research Institute for the Semi-Arid Tropics
C. R. Black
University of Nottingham, School of Agriculture
Abstract Modification of microclimate is one facet of agroforestry where potential exists for favourable interspecific interaction, but where few data are available. Studies of the possible beneficial microclimate modifications in agroforestry systems may usefully draw upon the detailed data already amassed for sole crops and some multiple cropping systems. Data are presented from rain-fed experiments carried out at ICRISAT (Hyderabad, India) during the 1985 and 1986 rainy seasons. In both years millet (BK 560) and groundnut (R33-1) were grown as sole crops and as an intercrop with a row ratio of 1:3 (millet:groundnut) and row spacing of 30cm. Instrumentation was installed in sole and intercrop plots. In 1986 millet (BK560) was grown as an alley crop between one year old Leucaena leucocephala hedges. The hedges were spaced at 3.4m intervals in sole leucaena and alley crop plots while millet row spacing, within the alleys and in sole plots was 47cm. Instrumentation was installed in sole and alley crop plots. In each experiment light interception, wind speed, saturation deficit, soil and leaf temperatures were routinely monitored. The microclimatic characteristics and their effects on yield in the two systems are compared. Additional data are presented from the agroforestry trial indicating how competitive interactions below ground may negate any beneficial changes in microclimate.
Many workers have attributed to agroforestry a capacity for the beneficial modification of microclimates (Huxley 1983; Brunig and Sander 1983). Microclimates have already been studied in some systems such as shelter-belt plantings (Guyot, this volume, Section 7) but measurements in systems containing both trees and crops are rare. At ICRISAT, studies on microclimatic modifications in conventional intercropping systems have been in progress for several years (Marshall and Willey 1983). The investigation of microclimates in agroforestry systems was a logical extension of this research. Monteith et al. (1981) observed that limitations in the study of microclimate often rest not in the collection of data but in the choice of appropriate forms of analysis. It is important not only to quantify changes in microclimate but also to relate them to plant responses. In order to predict the effect of changes in one component of the microclimate on the rate of specific plant processes, it is frequently necessary to resort to data already collected in controlled environment studies. In this paper we present results from rain-fed stands at ICRISAT (Hyderabad, India) during the 1985 and 1986 rainy seasons which allow comparisons to be drawn between intercropping and alley- cropping systems. Although these experiments were not specifically designed for this purpose, they demonstrate similarities between the two systems and serve to illustrate methods of analysis which relate microclimatic variables to growth and development.
Temperature and thermal time In order to describe the response to temperature of crop developmental processes many workers have used the concept of thermal time or accumulated temperature (e.g. Gallagher (1979) for wheat; Garcia-Huidobro et al. (1982a,b) for pearl millet; Leong and Ong (1983) for groundnut). This concept can be illustrated using the example of leaf appearance in pearl millet. The rate of development may be expressed as the reciprocal of the time (t) between the emergence of successive leaves from the whorl of the preceding leaf. This rate (leaves per day) is a linear function of temperature given by: 1/t = (T-Tb)/t [1] where temperature (T) exceeds the base temperature (Tb), below which no development occurs, but is less than the optimum temperature (To), above which increasing temperature reduces the rate of development. The The thermal time for the process,T, can be expressed as day degrees (C d) per leaf. Ong and Monteith (1985) have provided evidence that Tb and To are conservative over many developmental processes for a particular variety. Thermal time may be calculated from hourly or daily mean temperatures, although the latter should only be used in cases where T never exceeds To during the diurnal cycle. In the tropics, temperatures at mid-day often exceed To, in which case: l/t = [(t1/t)(T1-Tb) /T1] + [T2/t)(Tm-T2)/t2] [2] (Garcia-Huidobro et al. 1982b), where some fraction of the process is completed at a mean temperature T1 below To and in a time T1 and the remaining fraction is completed at a mean temperature T2 above To and in a time T2. Tm is the maximum temperature beyond which the rate of development is zero. ATI represents the degree-days above Tb, and T2 the day-degrees below Tm necessary for completion of the process under consideration. In our analysis accumulated thermal time during different periods of growth has been calculated from the equation: t = t1(TTb) +t2 (Tm-T2) t1/T2 [3] If the cardinal temperatures for different processes are similar, the T values may differ from process to process but the ratio t1/ t2 will remain constant.
Saturation deficit is a frequently measured microclimatic variable because of its importance in determining potential evaporation and plant water-use-efficiency. However it is a difficult quantity to relate directly to plant growth and development owing to its linkage with rainfall, temperature and soil moisture. Monteith (1986) used a relation between photosynthetic rate and vapour pressure deficit developed from laboratory studies (El Sharkawy et al. 1984). In this analysis actual net carbon exchange (N) is related to actual vapour pressure deficit (D) by the linear relation: N = Nm[ (l-(D-Do)/(Dm-Do) ] for D > Do and N = Nm for D £ D0 [4] where Nm is the maximum net carbon exchange, Do is the vapour pressure deficit below which N = Nm and Dm is the deficit at which net carbon exchange becomes zero. In our analysis we have calculated a saturation deficit factor, Z, from the equation: Z = l-[(D-Do)/(Dm-Do)] [5] (Monteith 1986, but with D = saturation deficit) for evaluating the possible effects of changes in saturation deficit on growth. It is assumed for this purpose that all other microclimatic factors remain constant, especially leaf temperature and soil moisture. If the microclimate was modified so that mean canopy temperature increased while saturation deficit decreased, the vapour concentration gradient between leaf and air might increase, decrease or remain constant depending on the balance between these two quantities.
In the 1985 and 1986 rainy seasons at ICRISAT (Hyderabad, India) pearl millet (Pennisetum americanum (L.)) Leeke var. BK 560) and groundnut (Arachis hypogaea L. var. Robut 33-1) were grown as sole crops and as an intercrop with a row ratio of 1:3 (pearl millet: groundnut) and a row spacing of 30 cm. In the 1986 rainy season, pearl millet (BK 560) was grown as an alley crop between hedges of one-year-old leucaena (Leucaena leucocephala (Lam.) de Wit). The hedges consisted of a double row of leucaena with 60 cm between the two rows and 25 cm between trees within each row. These hedges were spaced at 3.4 m in sole leucaena and alley-crop plots, while the pearl millet row spacing was 47 cm within the alleys (5 rows in each alley) and in the sole plots. At harvest, crops were sampled row by row and dry matter (DM) yields calculated on a 'row area' basis from the equation:
Grain yields were calculated in a similar manner. In each experiment the length of row sampled was 6 m. In the alley-cropping experiment the hedges were cut to 70 cm prior to sowing the millet, at 30 days after millet sowing, and again after millet harvest. At each cutting, 28 sample trees were measured for dry matter production in prunings and as dry matter increment in stems. Stem dry matter increment was calculated using an experimentally-determined regression of stem dry weight on stem volume and field measurements of stem volume. Tables 1 and 2 summarize the details of the intercropping and alley-cropping experiments, respectively.
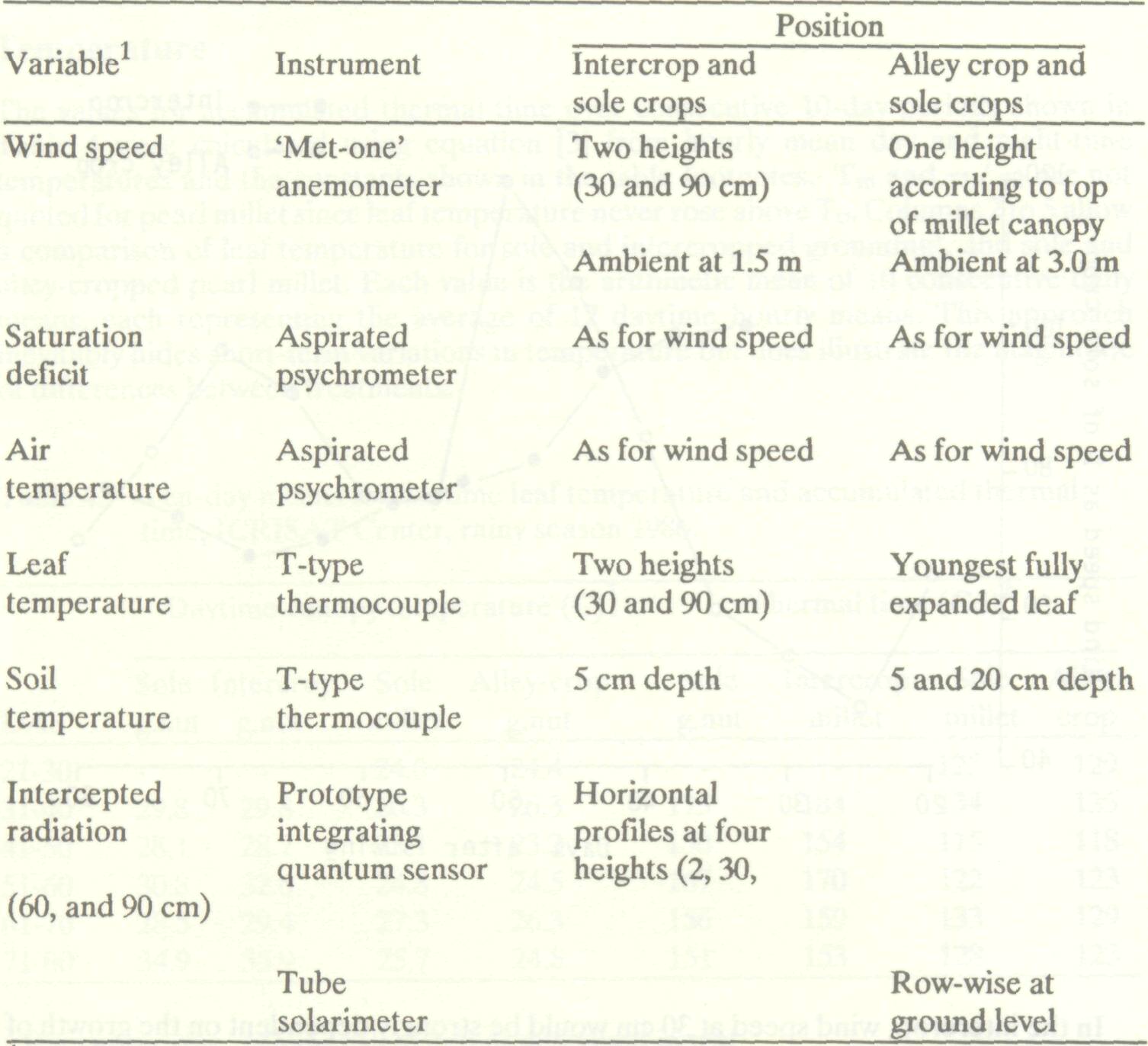
In each experiment wind speed, saturation deficit, leaf temperature, soil temperature and light interception were monitored. The variables measured are listed in Table 3 together with details of the instruments used and their positioning within the experiment. Signals from all instruments except the prototype integrating quantum sensor were scanned every 6 minutes by a data logger which computed and stored hourly averages. The prototype integrating quantum sensor was used in conjunction with a polycorder (Matthews et al. 1987) and measurements were made at midday once a week. Table 3. Instrumentation in intercropping and alley cropping experiments, ICRISAT Center, rainy seasons 1985 and 1986.
Although results from both the 1985 and 1986 intercropping experiments will be presented we will concentrate on a comparison of the 1986 intercropping and alley-cropping experiments.
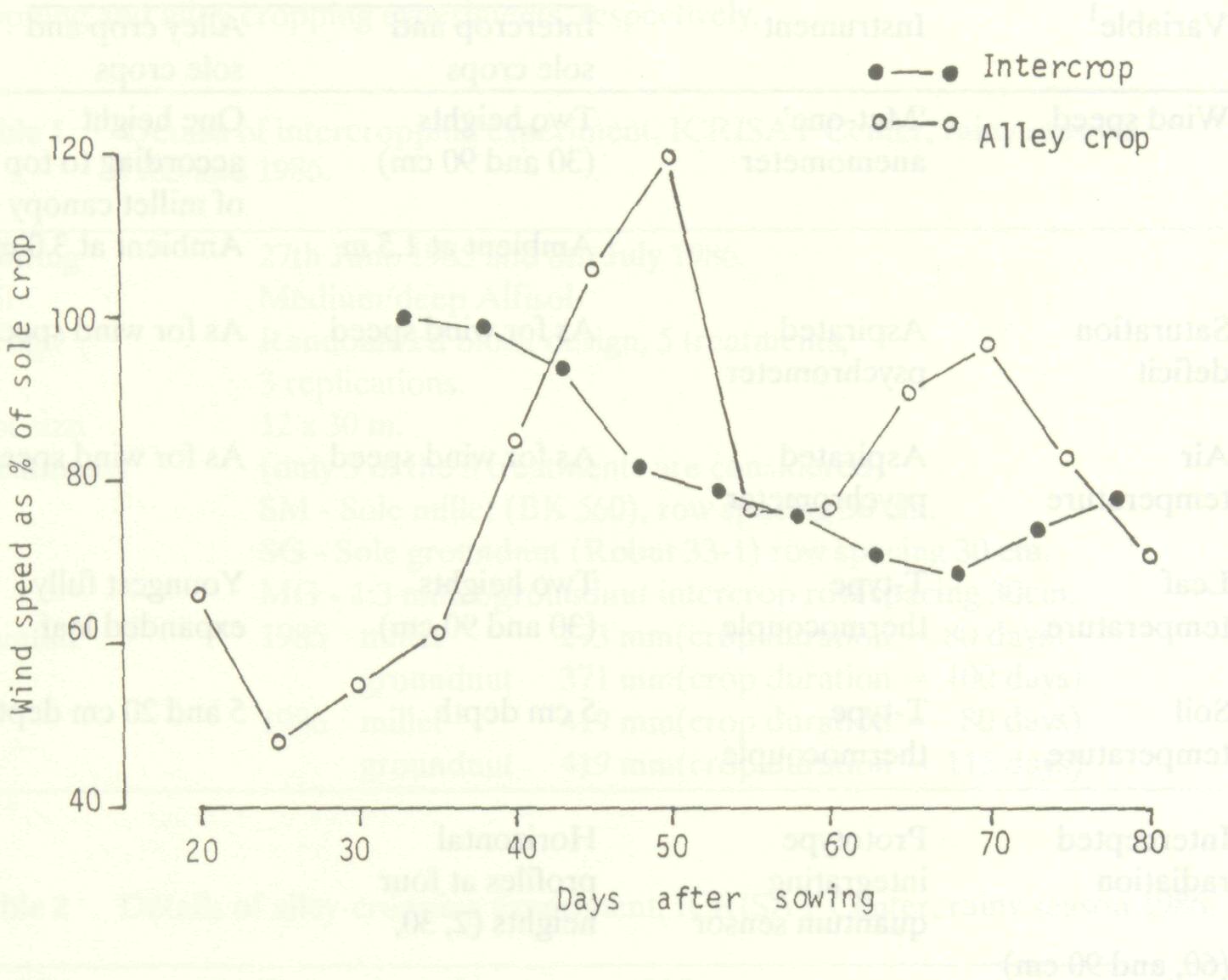
Figure 1 shows wind speed above the intercropped groundnut and alley-cropped pearl millet, expressed as a percentage of the corresponding sole-crop value. Wind speed above the groundnut intercrop (anemometer at 30 cm) was apparently unaffected by the pearl millet until almost 35 days after sowing (DAS), by which time the pearl millet had grown sufficiently to alter the pattern of wind flow over the groundnut. This effect became larger as the pearl millet grew in height and increased in leaf area. The difference in wind speed between the sole crop and the intercrop decreased after about 68 DAS as the pearl millet began to senesce.
Despite these problems, an important difference between the two systems was still evident. The intercropped groundnut experienced reductions in wind speed only during the last 40 days of millet growth and this to a maximum of 30%. In contrast, leucaena as a perennial reduced wind speed by up to 55% during the early growth of the alley crop, a feature which could be maintained throughout the pearl millet growing season with appropriate hedge management.
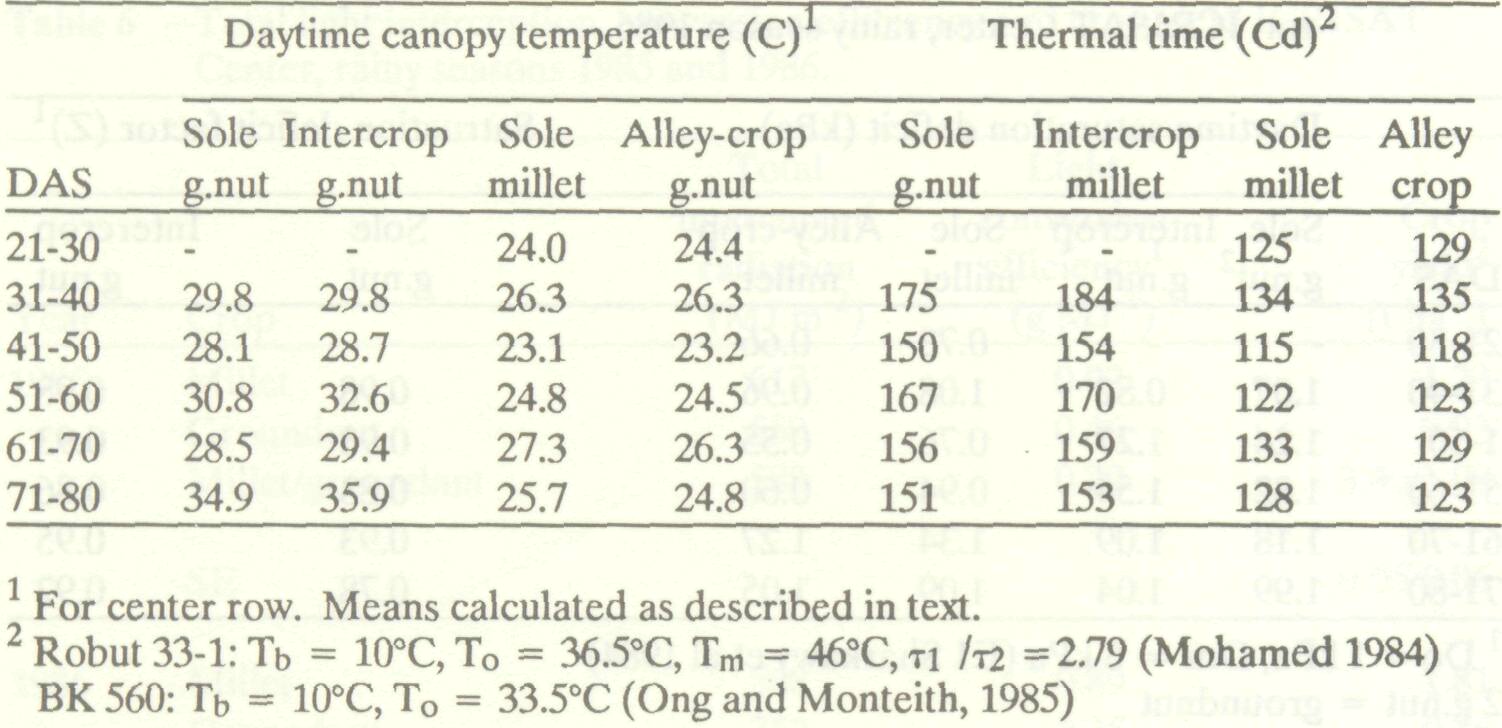
The values for accumulated thermal time over consecutive 10-day periods shown in Table 4 were calculated using equation [3] from hourly mean day and night-time temperatures and the constants shown in the table footnotes. Tm and t1 /t2 are not quoted for pearl millet since leaf temperature never rose above To. Columns 2 to 5 allow a comparison of leaf temperature for sole and intercropped groundnut, and sole and alley-cropped pearl millet. Each value is the arithmetic mean of 10 consecutive daily means, each representing the average of 12 daytime hourly means. This approach inevitably hides short-term variations in temperature but does illustrate the magnitude of differences between treatments. Table 4. Ten-day means for daytime leaf temperature and accumulated thermal time, ICRISAT Center, rainy season 1986.
Intercropping increased the 10-day mean daytime canopy temperature of groundnut by up to 1.8°C. This effect probably resulted from an increase in aerodynamic resistance, which would have reduced the ability of leaves to dissipate absorbed radiant energy by sensible or latent heat flux. Alternatively, competition for soil moisture in the intercrop might have reduced the amount of water available to the groundnut for evaporative cooling. Temperature differences between treatments were, however, small and amounted to only 21 degree-days from 31 to 80 DAS. A 1°C increase maintained over this entire period would have given a 50 degree-day difference in accumulated thermal time, sufficient to cause a difference in overall development of at most two days. The pattern in the alley crop was more complicated, but the differences between treatments were again relatively small. Canopy temperature in the alley-cropped pearl millet was similar to or slightly higher than that of the sole pearl millet between 21 and 50 DAS, as is reflected by the accumulation of 8 more degree-days during this period. Between 51 and 80 DAS, canopy temperatures were 0.3-1 °C higher in sole pearl millet than in the alley crop and the former accumulated 8 more degree-days. It thus appears that during this period shading by the leucaena may have reduced radiant energy fluxes to the pearl millet canopy to more than offset any decrease in the rate of heat loss caused by reductions in wind speed.
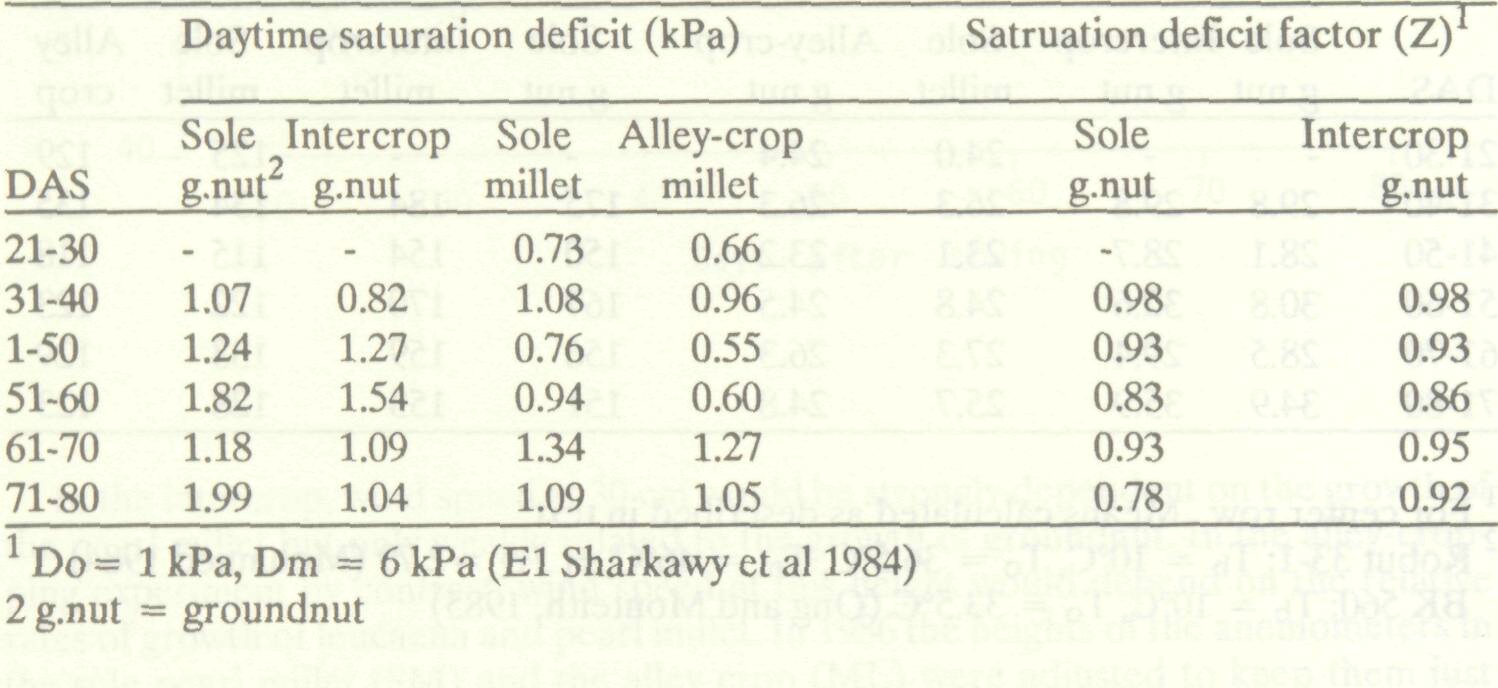
Table 5 shows mean daytime saturation deficits and calculated saturation deficit factors (Z) for consecutive 10-day periods in both intercropping and alley-cropping systems. The records suggest that groundnut experienced a lower saturation deficit when grown as an intercrop, with the difference rising to almost 1 kPa towards the end of the pearl millet season. This difference can be attributed to higher aerodynamic resistances within the intercrop canopy which would decrease vertical water vapour fluxes within and above the groundnut canopy. The extent to which these differences might affect crop growth (other factors remaining constant) is indicated by Z as shown in Table 5. These values were calculated from equation [ 5 ] on an hourly basis using the values for D0 and Dm determined by El-Sharkawy et al. (1984) for cassava. More rigorous analysis would obviously require the determination of D0 and Dm for groundnut. Daily means (12 hours) and 10-day means were then calculated from the hourly values. It is interesting to note that between 31 and 50 DAS the differences between treatments in mean saturation deficit were not reflected by the factor values. This results from our assumption that net carbon assimilation is reduced only when D is greater than D0and that there cannot be any increase in assimilation above the maximum rate if D falls below D0. Thus, changes in D below D0are of no importance in determining carbon assimilation, but may affect transpiration rate. Dry matter accumulation between 40 and 80 DAS would have been 89% and 93% of the maximum possible in the sole crop and the intercrop respectively if all other factors had been non-limiting. Table 5. Ten-day means for daytime saturation deficit and the saturation deficit factor, ICRISAT Center, rainy season 1986.
Saturation deficits were generally lower in the alley-cropping trial, and the millet experienced lower deficits as an alley crop than as a sole crop. Since the alley-cropping trial was planted 14 days before the intercropping experiment the data for specific 10-day periods are not directly comparable. We have not calculated Z for the alley-cropping experiment. As the deficits were generally close to or below D0 the small differences between the two treatments are unlikely to have significantly affected dry matter accumulation.
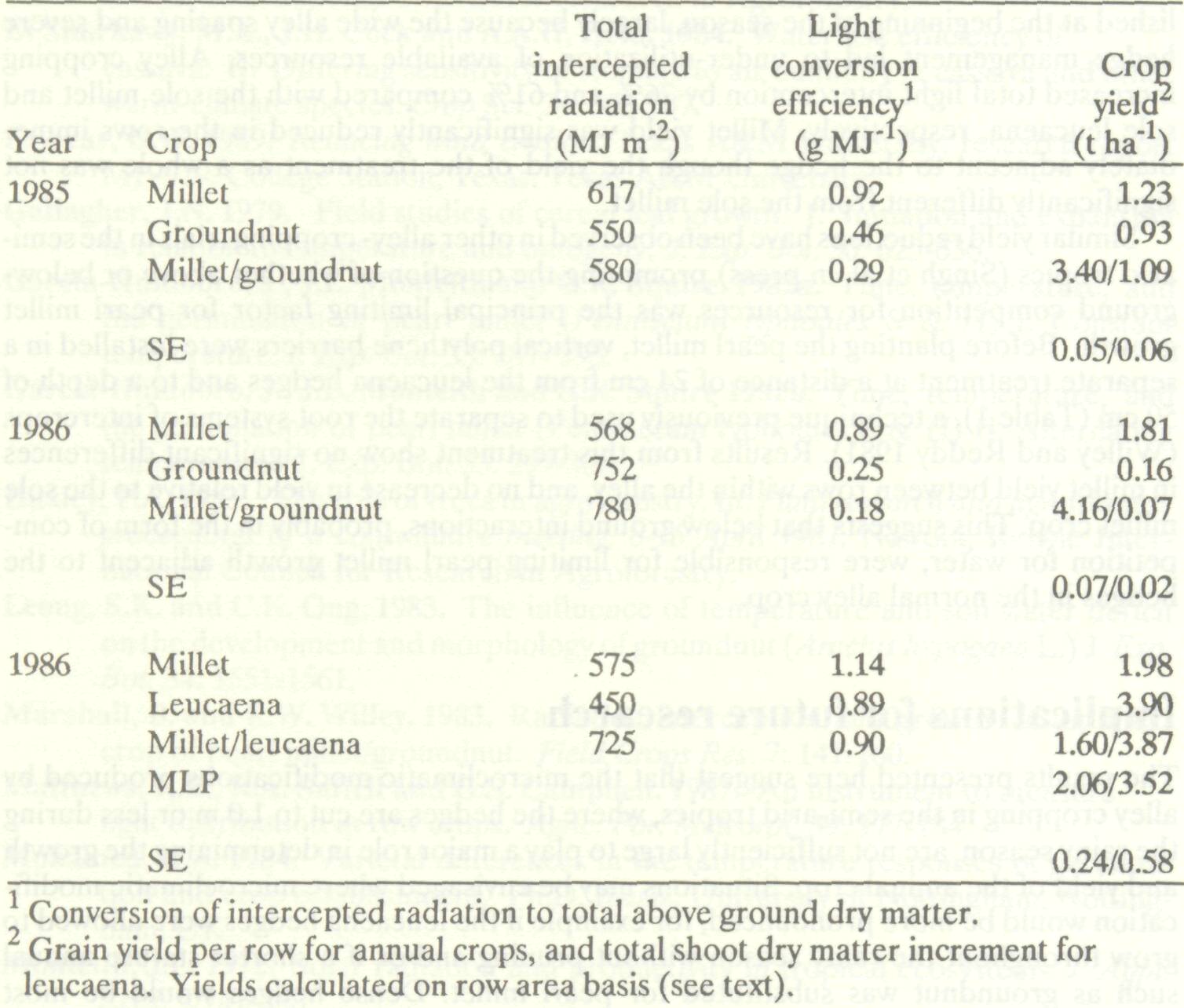
Table 6 shows total light interception, conversion efficiency, and yield on a row area basis for two seasons of intercropping and one of alley cropping. As dry matter accumulation is often considered to be proportional to accumulated light interception (Monteith 1972), changes in total light interception may be expected to affect yield. However, conversion efficiency, that is the quantity of dry matter produced per unit of intercepted radiation, is equally important. For instance Marshall and Willey (1983) and Willey et al. (1986) described the partitioning and utilization of light in a 1:3 pearl millet/groundnut intercrop and showed how the yield advantage of the intercrop could be attributed to increased light interception (per row) by the pearl millet and an increased efficiency of conversion by the groundnut. Table 6 Total light interception, conversion efficiency and crop yield ICRISAT Center, rainy seasons 1985 and 1986.
In the 1985 intercropping trial, yield of both species (on a row basis) was improved when they were grown as an intercrop, but in 1986 groundnut yield was very poor in the sole crop and even worse in the intercrop. These results cannot be explained by differences in light interception since groundnut intercepted more light in 1986 than in 1985. With respect to efficiency, pearl millet and groundnut performed substantially below their potential (Monteith 1972; Marshall and Willey 1983) in both 1985 and 1986. In both years, rainfall was well below the expected 600 mm during the groundnut growing season. The crops would therefore have been under drought stress for at least part of their lives, causing stomatal closure and reducing their ability to convert intercepted light energy into dry matter. The difference in crop performance between the two years originated largely from differences in rainfall distribution. Rainfall during the period of millet growth was more in 1986 than in 1985 but in 1986 no rain fell after the millet was harvested (Table 1), causing the groundnut to become highly drought stressed during the sensitive pod-filling stage. The yield difference between sole and intercropped groundnut in 1986 may be due to there being less water left in the intercrop profile after pearl millet harvest. In the alley-cropping system, rainfall was adequate during pearl millet growth except for a 15-day period soon after emergence. Total light interception and conversion efficiency for sole pearl millet were similar to those obtained in the intercropping experiment. Interception by leucaena was small even though the trees were already well established at the beginning of the season, largely because the wide alley spacing and severe hedge management led to under-utilization of available resources. Alley cropping increased total light interception by 26% and 61% compared with the sole millet and sole leucaena, respectively. Millet yield was significantly reduced in the rows immediately adjacent to the hedge though the yield of the treatment as a whole was not significantly different from the sole millet. Similar yield reductions have been observed in other alley- cropping trials in the semi-arid tropics (Singh et al. in press) prompting the question of whether above or below-ground competition for resources was the principal limiting factor for pearl millet growth. Before planting the pearl millet, vertical polythene barriers were installed in a separate treatment at a distance of 24 cm from the leucaena hedges and to a depth of 50 cm (Table 1), a technique previously used to separate the root systems of intercrops (Willey and Reddy 1981). Results from this treatment show no significant differences in millet yield between rows within the alley, and no decrease in yield relative to the sole millet crop. This suggests that below ground interactions, probably in the form of competition for water, were responsible for limiting pearl millet growth adjacent to the hedges in the normal alley crop.
The results presented here suggest that the microclimatic modifications produced by alley cropping in the semi-arid tropics, where the hedges are cut to 1.0 m or less during the rainy season, are not sufficiently large to play a major role in determining the growth and yield of the annual crop. Situations may be envisaged where microclimatic modification would be more pronounced, for example if the leucaena hedges were allowed to grow throughout the rainy season without pruning and/or if a shorter stature annual such as groundnut was substituted for pearl millet. Dense hedges would be most beneficial to crops in areas prone to wind damage (Fryrear 1969). However, in most cases competition for water would increase, probably to a degree that could not be compensated for by concurrent improvements in either light or water use efficiency. In the semi-arid tropics the prime consideration in the planning and management of agroforestry systems should be appropriate and efficient use of available water, with particular emphasis being placed on optimising its partitioning between tree and crop components. Whenever we seek to improve the aerial environment within specific systems we should always be aware of the possible implications and consequences of below-ground interactions.
This work was supported by the Isle of Man Board of Education (JEC) and research facilities were provided by the International Crops Research Institute for the Semi-Arid Tropics.
Brunig, E.F. and N. Sander. 1983. Ecosystem structure and functioning. In: Plant research in agroforestry: proceedings of a consultative meeting, 8-15 April. Nairobi, Kenya: International Council for Research in Agroforestry. El Sharkawy, M.A., J.H. Cock and A.A.H. Held. 1984. Water use efficiency of cassava. II. Differing sensitivity of stomata to air humidity in cassava and other warm-climate species. Crop Sci. 24:503-507. Fryrear, D.W. 1969. Reducing wind erosion. Texas A&M University, research report MP-929. College Station, Texas: Texas A&M University. Gallagher, J.N. 1979. Field studies of cereal leaf growth. I. Initiation and expansion in relation to temperature and ontogeny.J. Exp. Bot. 30: 625-636. Garcia-Huidobro, J., J.L. Monteith and G.R. Squire. 1982a. Time, temperature, and the germination of pearl millet (Pennisetum typhoides S & H) I. Constant temperature. J. Exp. Bot. 33: 287- 295. Garcia-Huidobro, J., J.L. Monteith and G.R. Squire 1982b. Time, temperature, and the germination of pearl millet (Pennisetum typhoides S & H) II. Alternating temperature. J. Exp. Bot. 33: 297-302. Huxley, PA. 1983. The role of trees in agroforestry. In: Plant research and agroforestry: proceedings of a consultative meeting, 8-15 April 1981. Nairobi, Kenya: International Council for Research in Agroforestry. Leong, S.K. and C.K. Ong. 1983. The influence of temperature and soil water deficit on the development and morphology of groundnut (Arachis hypogaea L.) J. Exp. Bot. 34:1551-1561. Marshall, B. and R.W. Willey. 1983. Radiation interception and growth in an intercrop of pearl millet/groundnut. Field Crops Res. 7:141-160. Matthews, R.B., R.A. Saffell and G.S. Campbell. 1987. An instrument to measure light distribution in row crops. Agric. For. Meteorol. 39:177-184. Mohamed, H.A. 1984. Varietal differences in the temperature responses of germination and crop establishment. Ph.D. thesis, University of Nottingham, Nottingham, UK. Monteith, J.L. 1972. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol. 9:747-766. Monteith, J.L. 1986. Significance of the coupling between saturation vapour pressure deficit and rainfall in monsoon climates. Exp. Agric. 22:329-338. Monteith, J.L., PJ. Gregory, B. Marshall, C.K. Ong, R.A. Saftell and G.R. Squire. 1981. Physical measurements in crop physiology. I. Growth and gas exchange. Exp. Agric. 17:113-126. Ong, C.K. and J.L. Monteith. 1985. Responses of pearl millet to light and temperature. Field Crops Res. 11:141-160. Singh, R.P., RJ. Van Den Beldt, D. Hocking and G.R. Kowar. In press. Alley cropping in the semi-arid regions of India. Paper presented at Alley Cropping Workshop, 10-14 March 1986, IITA, Ibadan, Nigeria. Wiliey, R.W. and M.S. Singh. 1981. A field technique for separating above- and below -ground interaction in intercropping; an experiment with pearl millet/ groundnut. Exp. Agric. 17: 257-264. Willey, R.W., M. Natarajan, M.S. Reddy and M.R. Rao. 1986. Cropping systems with groundnut: resource use and productivity. In: Agrometeorology of groundnut: proceedings of an international symposium, 21-26 August 1986. Niamey, Niger: International Crops Research Institute for the Semi-arid Tropics.
*Submitted as ICRISAT Conference Paper No. 362. |