![]()
An e-publication by the World Agroforestry Centre
METEOROLOGY AND AGROFORESTRY

|
|
An e-publication by the World Agroforestry Centre |
|
METEOROLOGY AND AGROFORESTRY |
|
|
SECTION 2 : Basic principles Climate and plant responses: the use of phenological data to help with the choice of woody species for agroforestry systems PA. Huxley, E. Akunda and A. Repollo*
International Council for Research in Agroforestry (ICRAF) Abstract The value of interpreting a woody plant's behavioural characteristics in relation to environmental changes, both aerial and edaphic, is briefly discussed in relation to under-standing species adaptability. Possibilities for managing the external environment and/or the plant itself, so as to synchronize vegetative growth and flowering/fruiting patterns favourably in relation to expected climatic sequences, can also be an outcome of such studies. The methods of investigating phenological behaviour are outlined, and two examples are given from a preliminary branch phenology study carried out at ICRAF's Field Station to illustrate the results of such investigations. Introduction Plant phenology is the study of changes in the plant's behavioural characteristics in relation to environment. There is an extensive literature on this subject (e.g., Lieth 1974; Huxley 1983), including many papers relating to the phenology of tropical woody species (e.g., Huxley and Van Eck 1974). The practical value of understanding the relations between climate and crop growth phases has been discussed, with examples, by Watts Padwick (1979), and the 'heat unit' or day-degree summation approach to predicting crop plant responses has also been extensively reported in the literature (e.g., Iwata 1975). Although many of the comments that follow apply to plants in general, the present paper is specifically concerned with the phenology of woody perennials. Phenological studies on areas of vegetation can present problems because individual woody plants can sometimes be out of phase with others of the same species, depending on the influences imposed by the site, other external influences and by internal rhythms. Large samples are often required, therefore, for the purpose of describing the behaviour of vegetation complexes (Grouzis and Sicot 1981). However, providing the limitations of the method are recognized, recording behavioural responses and climatic sequences for just a few specimens of any woody species can certainly be informative, as long as the interaction with site characteristics can be clearly defined. Such an activity can provide information on adaptability of woody species, as well as clues suitable plant management operations. This type of information can help plan field trials, especially for newly-introduced exotics and/or indigenous species that are being tried in new regions. This paper first briefly discusses these aspects, then describes the simple methodologies that can be used to obtain phenological information from the field, finishing with two examples of records that have been obtained from a preliminary study at ICRAFs Machakos field station.
Plant strategies Plant species possess a very wide range of strategies to help them establish their place in particular ecological niches (or clines). For well-adapted species the necessary steps in growth and development ('phenophases') have become adjusted and properly sequenced both to seasonal changes in climate, and to influences such as the phenology of associated species competing for a place in the same niche. At the extremes individual species can be found either confined to a highly specific ecological situation or they may traverse a wide range, obviously, the factors underlying the distribution of a plant species in space can be extremely complex. The ultimate control of morphological and physiological characteristics that enable plants either to be best-suited to a specific niche, or to spread to others, depends upon their genetic 'fitness' or 'flexibility". However, even species that are restricted location-ally will be so confined through some particular set of evolutionary circumstances; yet they may still possess a useful degree of morphological and/or physiological plasticity. This is an underlying feature that needs to be borne in mind in species selection procedures. A comparison of the growth and development patterns as expressed by any woody species in different environments is, clearly, a valuable procedure to provide information on such matters.
When we say that a woody species is 'adapted' to a particular environmental situation we mean that: (a) the range of soil conditions under which it finds itself is, for each and every soil factor that affects the plant, within the limits that encourage normal growth and development; (b) there are no climatic extremes that will eliminate the species; and (c) the probability of obtaining a suitable length and sequence of climatic conditions for growth and development processes in any one season is high. The absence of other detrimental biotic factors (e.g., pests and diseases organisms) is also important, of course. Assuming that soil conditions encourage growth, then plants that are adapted to a particular climate will progress through their various phenophases in a regular and unimpeded way (Huxley 1983). Species will vary in these patterns of response, perhaps quite markedly, but each can be expected to behave according to the particular strategy inherent in the pattern originally established at its centre of origin. Thus regulatory controls will be triggered by appropriate climatic conditions to result in leaf initiation, bud development and leaf expansion at an appropriate part of the season; and similarly for flower initiation, anthesis, pollination, fertilization and fruit growth and maturation. Growth cessation, if it occurs, will similarly be brought about by a combination of internal (endogenous) and external (exogenous) factors. The climatic conditions during each phenophase will also need to be such as to maintain all the different physiological processes at a balanced rate.
If a woody species is ill-adapted, then it is usually clear at an early stage that the sequence of phenophases is not properly sequenced or entrained to climatic changes, either (a) because what might be expected from its normal behaviour at its origin does not occur or (b) it behaves in a way that suggests that it will not be able to make good use of the climatic pattern in which it now finds itself. Of course, in the latter case, it may be possible, by using suitable management practices (e.g., lopping or pruning), to adjust the sequence of phenophases to a more suitable entrainment, and the phenological information gained by studying its behaviour in the field will provide information about the best times at which to make such management interventions. In these circumstances, there may be a case for recording the phenological behaviour of the woody species as modified and conditioned by a set or sequence of manipulations (lopping at different parts of the season), or to obtain some clues about this through plant or branch manipulation on individual plants. This is discussed further below.
Management of the external environment Management practices which most obviously effect plant phenology are those that impose, or alternatively relieve some degree or another of environmental stress. This is often done by manipulating soil factors. For example, as with climatic factors, soil conditions can enhance, slow or interrupt the proper sequence of phenophases; in this case by modifying water and/or nutrient supply. In temperate regions the application of nitrogen fertilizers to apple trees in the autumn in order to affect flowering next spring, is an interesting example of this. Additionally, detrimental soil conditions such as toxic effects of excess aluminium/manganese, or heavy metals, or waterlogging markedly affect the phenology of woody species. With trees/shrubs we are, however, much more limited than with high-value annual crop plants in the ways that we can attempt to modify the soil environment in order to manipulate plant behaviour for production purposes. The most feasible way is through site selection. The choice of soil management practices is often limited although, in agroforestry, we can add or retain plant residues and, perhaps, add some fertilizers. Where soil conditions operate to change the phenological pattern by prolonging the vegetative phase, as when there is added water or nitrogen, such 'surpluses' can sometimes act disavantageously with regard to the objectives for managed systems, particularly when they cause a delay in reaching the flowering/fruiting phase which may reduce yields. Irrigation can simulate rain and both enhance soil water status and, to some extent, improve aerial humidity profiles. It can thus initiate and/or prolong vegetative growth. There are many examples where 'sequence optimization' of the growth of different agricultural crops is achieved in this way. However, it is usually not feasible to use irrigation with multi-purpose trees in most agroforestry systems. Manipulation of environmental stress by controlling exposure is another way to modify phenological behaviour. Although subject to the same factors, woody species are usually used to create the stress-alleviating conditions for lower storey crops (e.g., as windbreaks, shade trees, etc.). Shading will also affect the sequence of phenophases (i.e., prolong the vegetative phase). The most likely situation in agroforestry is an increasing level of shade in the lower parts of a multi-storey canopy, or at a tree/tree interface.
Most of what has been discussed so far has emphasized the external controls over plant behaviour as imposed by either suitable or unsuitable climatic and soil conditions. Internal (endogenous) regulatory processes can be important also, and these can be modified to a greater or lesser extent by various forms of plant management. They are more feasible with trees and shrubs than attempts at climate or soil modification. For example, by far the most useful management procedure to consider for multipurpose trees is some form of removal of plant parts. This can be by (a) the removal of above-ground vegetative parts by pruning or lopping (the most usual practice); (b) the removal of flowers and fruits (usually done by removing also the vegetative parts on which flowering takes place); and (c) root removal by some form of trenching or other root pruning process. Each of these is briefly discussed below.
Pruning or lopping of aerial parts will affect the annual biomass increment produced (usually by reducing it according to the severity of lopping), and it will affect the subsequent distribution of dry matter. It will also act to modify the initiation, duration and cessation of the subsequent vegetative phenophases. These effects can vary very much both between species, and depending on the time at which the shoots are lopped. They may be promoted to occur sooner than normal, or they may be delayed. This will depend on the condition of the lateral buds, the level of stored materials in the shoot, and the responses engendered by cessation of any hormone supply from the removed shoot apex. Pruning/lopping is, therefore, a powerful management tool for multipurpose trees. However experiments to test the factors involved (time, intensity and frequency of lopping, type of plant part removed) must be rationally thought out in order to take advantage of the known patterns of climatic change at the experimental site, and their likely effects on the phasing of plant growth. Some preliminary information about the plant's phonological responses in its natural state will enable a much more precise approach to be taken to this form of experimentation which, otherwise, will remain as partial guesswork. Indeed, lopping times for experiments must be defined in terms, not of calender periods but of actual weather patterns and the plant's phenological response if they are to be meaningful.
Removing flowers, fruits or vegetative material on which flower initiation is known to occur will have two effects. The first effect is similar to that mentioned above for removing vegetative parts; the second is that of removing a future significant and powerful sink for assimilates and nutrients. When woody perennials produce fruit, the drain on resources stored or current can markedly affect the pattern of subseqent vegetative growth depending on the degree of fruiting experienced in relation to resources available. This, in turn, is affected by favourable or unfavourable climatic sequences during the fruit maturation period and, perhaps, by the way in which a previous favourable or unfavourable season has affected the plant's levels of stored nutrients and carbohydrate. In some cases biotic factors can also be involved (e.g., damage to the leaf canopy by pest and disease organisms after a fruit load has been established). It is not uncommon in the tropics to see two plants of the same species at the same site completely out of phase with one another because a prior set of factors brought about a prolific flowering in one but not in the other. The subsequent development of a heavy fruit load on the first specimen was, in these circumstances, sufficient to disrupt its expected sequence of phenophases. This may occur even to the point where it enters a bi-annual or bi-seasonal flowering/fruiting cycle or, in an extreme case, has flowering delayed for several seasons. The management of flowering in order to adjust fruiting levels is, therefore, another possible tool with which to manipulate climatic 'entrainment'.
Root pruning can modify subsequent phenology. In some cases (e.g., with some temperate fruits) root pruning is used to promote flowering when this is unduely delayed by an extended stage of vegetative growth in the early years. When this occurs it is probably a result of a change in the balance of plant hormones which are produced by roots (e.g., cytokinins and gibberellins) and which regulate growth in other parts of the plant. Removing roots can certainly modify the subsequent extent of nutrient and water removal from the adjacent soil. The practice of root pruning is common in many agroforestry situations because tillage relatively close to trees or hedges can help restrict their competitive influence. There seems to be relatively little information about how root pruning may affect subsequent within-season phenophases of multipurpose tree species growing in the tropics and sub-tropics, but it could be a useful area to look into as a potentially useful management procedure.
Other management practices or situations that can influence the phenology of woody plants in the tropics are those that can indirectly affect stress patterns. For example, planting density, choice of site to affect exposure or shelter, aspect, slope (inasmuch as it affects run-off and soil water) and, of course, the association of woody perennials with crops and/or grasses. With what has been said so far, there seems little doubt that phenology records can usefully be included in all agroforestry field trials in order to establish information about the basic phenological sequences and to compare the effects of different treatments.
The methods for obtaining phenological records of above-ground sequences of plant behaviour can be extremely simple. It is, therefore, surprising that more use is not made of this approach. Obtaining information about patterns of fine-root growth are more difficult and time-consuming, although they may well be of considerable value.
Whenever phenological data are to be collected, concurrent meteorological information must be gathered either from the site itself or nearby. It should be remembered, in this respect, that topographical changes can bring about quite a large difference in local climate in the tropics, especially with regard to night temperature, humidity and dew point (Huxley and Beadle 1964). Such topoclimatic variation (especially night temperature differences) can affect the phenological progress of crop plants, although little is known with respect to tropical trees and shrubs. Bearing these points in mind we can collect phenological information in three ways.
A set of very useful information can be obtained simply by taking weekly or two-weekly sets of observations of whole tree behaviour. This should always be with undisturbed specimens growing in a representative site (the 'control'), but it can be extended to include similar trees in different kinds of sites, or under particular experimental conditions or management practices, if this is of interest. The observations can be made by a trained field assistant and they can be quickly and easily checked. An indication of the intensity of each factor being recorded can be obtained by scoring 1 to 5 (Huxley and Van Eck 1974). Observations would normally be to identify the periods over which the following occur:
Branch phenology A close scrutiny of a branch will often expose details of how it has grown over the past few seasons, as well as providing information about where and what kind of fruiting sites are formed (Figure 1). Making measurements on individual branches can be rather time-consuming so that this exercise has to be carefully thought out. Such a study can include some manipulation which may provide clues relating to tree management by lopping or browsing. Usually, a number of representative branches are chosen at the start on a random sample of about 3 5 trees of the species under investigation that are growing in the selected site conditions. Unless a large amount of replication is possible it will not be possible to make rigorous statistical comparisons, and any replication is just to obtain a better sample mean. If branches on individual trees do not behave similarly this, too, is useful information. Only large differences in phenological patterns will emerge from this kind of investigation, but at least they can be presented as a tabular or graphical record.
Once selected, branches are marked with a paint spot at a suitable internode, not too far from the apex but far enough back to be sure that the internode is mature. It is important to use a non- toxic paint, such as a water-soluble plastic emulsion, making certain that it is not placed on green, tender tissue. Usually the mark can be made just below the 7th or 8th node from the shoot apex. The branch should be labelled to indicate its number and/or location in the tree and any subsequent treatment that is to be imposed, using a label that will persist in a reasonable state over several seasons. The label must be secured in a way that will not damage the shoot. We have found it useful to attach an extra length of coloured string or twine so that each label can be quickly found. The datum spot can be moved after a branch has grown to a point where it involves too much work to count the nodes and/or it exceeds the length of the ruler. It is necessary to note in the records the point to which it has been re-located, and to measure and record both the old and new lengths. One set of branches should be maintained as a control, with treatments imposed on other sets. For example, the removal of the shoot apex at different parts of the season may provide clues as to the species' potential for re-sprouting at these periods. This approach has to be treated cautiously as the growth regulatory mechanisms of the treated branches may be over-ridden or compensated for by adjacent ones. Thus a positive response of a single shoot to apex removal will indicate that the whole tree or bush is likely to respond in the same way at that part of the seasons. However, a negative response may indicate that growth of the branch still remains suppressed, and a more severe lopping treatment of the whole plant will be needed to test its potential for re-growth. The same applies if flower or fruit-removal is to be tested as a method of changing the plant's phenological entrainment. Records should be taken at sequential intervals, the actual time being related to periods of climatic change rather than calender dates, as follows:
There is obviously a limited period during which branch phenological data can be taken before the structure becomes too complicated to record easily. The method is therefore best-suited to investigations that are limited to just a few seasons.
As the study of an individual woody species progresses it may become necessary to understand more about the way the plant's development sequences relate to climatic and/or management factors, especially the progress of leaf initiation over time (the plastachron), and the appearance of expanding leaves with time (the phyllochron) (Figure 1). These two may or may not be closely correlated depending on whether a period of vegetative bud dormancy occurs; but they are both key stages strongly affected by climatic factors, e.g., by available soil water and atmospheric humidity (or both), and by ambient temperatures. Although results of such a detailed investigation can help with the interpretation of the plant's responses to management, it is essential first to decide whether or not this level of detail is really required. Phyllochron records will be obtained from branch phenology studies (see above), but the plastochron can only be defined by dissection of vegetative apices sequentially with time. This is not difficult and it can be done with a high-powered binocular microscope, taking care to sample representative shoot apices. Similarly, a knowledge of the time at which flower or inflorescence buds are initiated can be essential to the proper understanding of the way seasonal climatic variations affect subsequent flowering and fruiting. Woody plant species differ considerably in where (i.e., on what part of the branch) and when flower initiation take places (Huxley 1985). It can be in a previous season, or only just a short time before anthesis (flower opening). If knowledge of the effects of climatic or management factors on flower initiation is required, samples of appropriate buds should be dissected. Buds that have progressed from the vegetative to the developmental stage can nearly always be detected because their morphology changes, and the primordial shoot apex becomes flattened with flowering initials becoming evident. A prior investigation of this formative bud and development process has to precede the main investigation if such morphological changes have not been previously characterized for the species under test. Again, this is not difficult or particularly time-consuming.
Methods for studying roots are described in several publications (e.g. Bohm, 1979; Schuurman and Goedewaagen,1971.) All of them are time-consuming and laborious and require special facilities or equipment. For most phenological studies, patterns of below-ground activity may be as important (although much less easily observed) as above-ground activity. Indeed, it is to be expected that tropical woody perennials have as wide range of below-ground root growth strategies (for fine roots, at least) as they do for above-ground leaf production. Furthermore, the ecological success of many tropical woody species may well depend, at least in part, on the differential phasing of root and shoot growth if they are to compete successfully with associated species of one kind or another (see Huxley and Turk 1975 for an example of root-shoot growth phasing). It seems likely, therefore, that at some stage many investigators of multipurpose trees will wish need to incorporate a study of what is happening below-ground. Probably the simplest approach that can be suggested at present is to make a succession of observations and/or measurements of topsoil root activity by taking core samples and looking for the the presence of live, actively- extending fine roots. This procedure would initially be purely qualitative; but it can be made quantitive by washing, collecting and measuring the length of different root fractions (Ford and Deans 1977). At the same time the presence or absence of root nodules can be observed, if appropriate. The effectiveness of nodules can be defined quantitatively by estimating potential rates of di-nitrogen fixation, using the ethylene acetylene reduction technique. Such observations may provide useful clues as to whether or not a more detailed below-ground study is needed later.
We give here two examples from a preliminary branch phenology study on seven woody species that commenced in 1985 at ICRAF's field station at Machakos, Kenya. The objectives were to test the feasibility of this approach and to-assess its value for the practical assessment of woody species introduction and management. The methods used were as outlined in the section above and, although sample sizes were very small, the data generally coincide with the observable behaviour of all similar specimens in the plot. The reader is referred to Huxley and Van Eck (1974) for an example of a long-term study of single whole plant phenological observations on a number of woody species at one tropical site.
Growth of uncut lateral branches Figure 2 shows the periods of extension growth of typical primary lateral branches of young trees (4 years from planting out) of Psidium guajava (guava), Acacia stulmannii, and Leucaena leucocephala cv. K.8, over three rainy seasons from August 1985 to February 1987. Monthly rainfall totals are also shown. The first period of rains (short rains) started in October 1985 and continued through January 1986; the second (long rains) was from March to June 1986; and the third (short rains) was from October 1986 to January 1987. Intervening periods were virtually dry seasons. This bimodal pattern is normal in this region. The elongation growth of intact primaries (branch A) is shown as well as the continuation of primary growth by a secondary after removing the apex (branches C & D). Guava originated in the American tropics but is quite widely spread as a fruit crop through tropical and sub-tropical regions; it has been cultivated in various parts of Kenya for many years.A stulmannii is an indigenous species common to the drier areas of Kenya. Leucaena is native to Central and South America and is usually considered best adapted to higher areas with more than 1000 mm of annual rainfall (Brewbaker 1987). These three species exhibited quite different growth patterns of uncut primaries. Branches of A. stulmanii grew in length mainly during the rainy periods but ceased, as is common with many tropical tree species, prior to the end of the rains. However, in the short rains of 1986/87, a second phase of growth extended into the dry season. The most vigorous extension growth was in the short rains. A small amount of branch extension also occurred during the dry period in September. Guava showed a great deal of pre-rains extension growth of primaries in each of the three rainy periods, and extension again ceased (first two seasons), or slowed (last season) before the end of the rains. Branches of leucaena grew vigorously on these young trees well before the short rains started in October 1985 and again, briefly, post-short rains (February 1987). Very little extension growth occurred subsequently. In the last season shown, extension growth began, weakly, in the dry season well before the start of the short rains of 1986/87, and continued, in the same way, into the start of the next dry season (February 1987).
A. stulmanii flowers on the previous season's vegetative flush and, if a period of environmental stress sets in, may or may not set fruit and/or shed young fruits. If not cut back, guava flowers on short lateral shoots arising on maturing parts of a branch and/or on 'spurs' found in the axils of leaves on older wood. Flowering accompanies or follows a period of vegetative growth. Leucaena forms inflorescences on new-shoot, lateral branches arising near the apex of extending, primary shoots, i.e., on current season's growth. Its flowering was erratic and badly timed. The phenological patterns show that both guava and A. stulmanii are adapted to the environment at this site. There are no particular soil problems, apart from a generally low level of soil fertility on these Oxisols; and, at least from the three seasons of observations reported here, they could be seen to be suitably entrained to the climatic pattern. Leucaena presents an entirely different case, however. Flowering occurred only towards the end of the first short rains of 1985 (November). It co-incided with the early start of the long-rains of 1986 (February), but another flowering occurred at the end of these in May, and yet again in the following dry season (August). There appears to be no mechanism whereby leucaena can shed developing fruits, and the depletion of plant reserves during the fairly long fruit growth and maturation periods were not supported by vegetative flushes. A consequence was, eventually, a very weak and intermittent vegetative flush starting early in the pre-short rains of 1986, which continued into the February 1987 start of the dry season. The sequence shows clearly that Leucaena leucocephala is not inherently adapted to this site. It can, of course, be forceably entrained by a suitable lopping treatment to be somewhat better suited to it. This has been done at Machakos, so that its non-adaptability is unlikely to be a consequence of unsuitable soil (e.g., low pH) at this site.
In order to obtain some evidence on plant response some branches on each tree had 15cm or so of the apex removed at specific times in the season (see methods sections above). For all three species shown in Figure 2 it took approximately a year before apical growth on the primary was continued (Branches C and D) by a suitable (i.e. proximal) lateral shoot . Nor did it make much difference whether the primary apex had been removed prior to or during the short rains of 1985/86.
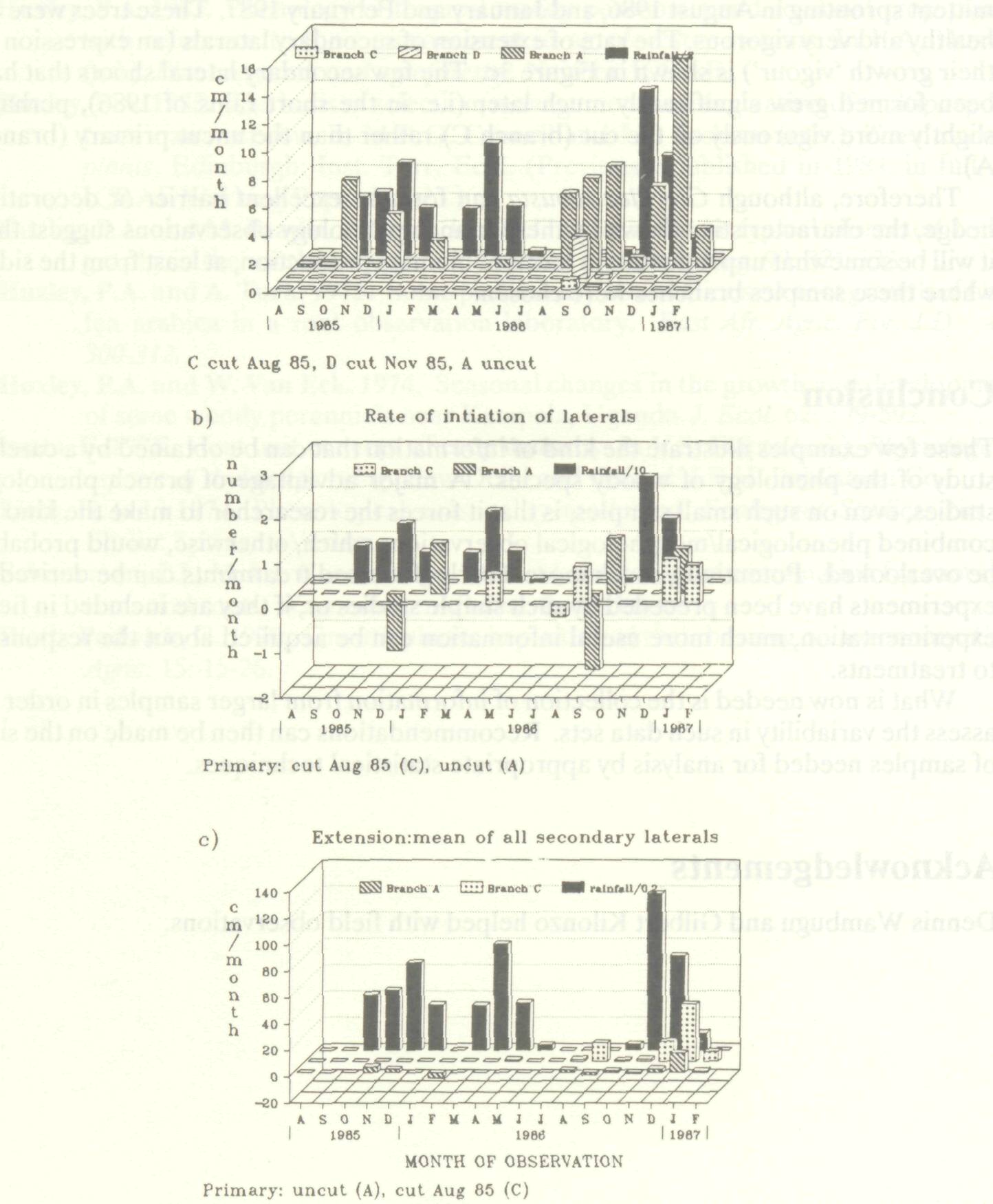
Figure 3 details the initiation and growth of secondary laterals of another well-adapted species, Grevillea robusta. The uncut primary lateral branches (branch A) grew vigorously in the first and third rainy seasons (Figure 3a). Primary branch extension almost completely coincided with the start of the short rains in October 1985. Some pre-long rains growth occurred in the dry February of 1986 (but there was still probably some soil water storage at depth then). For some reason, there was little extension of primary laterals in the long rains of 1986. Rapid extension started again well before the short rains beginning in November 1986, with another flush towards the end and into the dry season (cf. A stulmanii).
As with the other three species, there was a long delay in renewal of terminal growth on primaries once a primary shoot has been cut (branches C & D, Figure 3a). Counts of secondary lateral buds and their extension growth are also shown in Figure 3. Some young growing laterals (identified when longer than one centimeter) died back on the uncut primaries (branch A) in Grevillea in the short rains of 1985/86 (Figure 3b). A few more sprouted when these rains had ceased in January 1986. A similar pattern occurred prior to and at the start of the short rains of 1986/87. The example of a cut primary (e.g., in August 1985 - branch C) showed that nothing happened until April of the following year. There was little dieback of young laterals but some subsequent intermittent sprouting in August 1986, and January and February 1987. These trees were all healthy and very vigorous. The rate of extension of secondary laterals (an expression of their growth 'vigour') is shown in Figure 3c. The few secondary lateral shoots that had been formed grew significantly much later (i.e. in the short rains of 1986), perhaps slightly more vigorously on the cut (branch C) rather than the uncut primary (branch A). Therefore, although Grevillea robusta can form an excellent barrier or decorative hedge, the characteristics shown by these branch phenology observations suggest that it will be somewhat unproductive in terms of biomass production, at least from the sides where these samples branches were chosen.
These few examples illustrate the kind of information that can be obtained by a careful study of the phenology of woody species. A major advantage of branch phenology studies, even on such small samples, is that it forces the researcher to make the kind of combined phenological/morphological observations which, otherwise, would probably be overlooked. Potentially, much more clearly- focussed treatments can be derived if experiments have been preceded by such simple studies or, if they are included in field experimentation, much more useful information can be acquired about the responses to treatments. What is now needed is the collection of information from larger samples in order to assess the variability in such data sets. Recommendations can then be made on the size of samples needed for analysis by appropriate statistical techniques.
Dennis Wambugu and Gilbert Kilonzo helped with field observations.
Bohm, W. 1979. Methods of studying root systems. Ecological studies. Berlin: Sprin-ger-Verlag Brewbaker, J.L. 1987. Leucaena: a multipurpose tree genus for tropical agroforestry. In H.A. Steppler and P.K.R. Nair (eds.), Agroforestry: a decade of development Nairobi: ICRAF. Ford, E.D. and J.D. Deans. 1977. Growth of a Sitka spruce plantation: spatial distribution and seasonal fluctuations of lengths, weights and carbohydrate concentration of fine roots. Plant Soil 47: 463-485. Grouzis, M. and M. Sicot. 1980. A method for the phenological study of browse populations in the Sahel: the influences of some ecological factors. In H.N. Le Houerou (ed.), Browse in Africa, the current state of knowledge. Adddis Ababa: ILCA. Huxley, P.A. 1983. Phenology of tropical woody perennials and seasonal crop plants with reference to their management in agroforestry systems. In P.A. Huxley (ed.), Plant research and agroforestry. Nairobi: ICRAF. Huxley, PA. 1985. The basis of selection, management and evaluation of multipurpose trees - an overview. In M.R.G. Cannell and J.E. Jackson (eds.), Trees as crop plants. Edinburgh: Inst. Terr. Ecol. (Previously published in 1984, in full, as ICRAF Working Paper No. 25.) Huxley, P.A. and M. Beadle. 1964. A local climatic study in typical dissected topography in the southern region of Uganda. Meteorol. Mag. 93: 321-333. Huxley, PA. and A. Turk. 1975. Some preliminary observations on root growth of Coffea arabica in a root observation laboratory. East Afr. Agric. For. J.D 40: 300-312. Huxley, PA. and W. Van Eck. 1974. Seasonal changes in the growth and development of some woody perennials near Kampala, Uganda.J. Ecol. 62: 579-592. Iwata, F. 1975. Heat unit concept of crop maturity. In V.S. Gupta (ed.), Physiological aspects of dryland farming. New Delhi and Oxford: I.B.H. Publishing Co. Lieth, H. (ed.) 1974. Phenological seasonality modelling. Ecological Studies. No. 8. Berlin: Springer-Verlag. Schuurman, J J., MAJ. Goedewaage. 1971. Methods for the examination for root systems and roots. Wagenningen: Pudoc. 2nd edition. Watts Padwick, G. 1979. Growth phases in plants and their bearing on agronomy. Exp. Agric. 15:15-26.
* Professor A. Repollo Jr., DMMMSU, Bacnotan, La Union, The Philippines. Visiting Senior Fellow at ICRAF during 1985-86. |