An e-publication by the World Agroforestry Centre
AGROFORESTRY A DECADE OF DEVELOPMENT 

|
|
An e-publication by the World Agroforestry Centre |
|
AGROFORESTRY A DECADE OF DEVELOPMENT |
|
|
section 4 Chapter 13 Pedro A. Sanchez Introduction Agroforestry systems are generally perceived to be sustainable and to enhance soil properties. Growing trees in conjunction with annual crops or pastures is believed to provide a more thorough plant cover to protect the soil from erosion and a deeper or more prolific root system to enhance nutrient cycling. Shortly after its creation, ICRAF organized a state-of-the-art review of soils research and agroforestry (Mongi and Huxley, 1979). A dearth of available, solid research data about the impact of agroforestry on soils and vice versa was obvious at that time (Sanchez, 1979). Since then, soils research in the tropics has gained considerable momentum, as has soils research in agroforestry (Nair, 1984, 1987; Young, 1986 a, b, c, 1987; Young et al, 1986. The objective of this paper is to critically evaluate the various hypotheses that have been advanced on the soil-productivity and sustainability aspects of agroforestry and suggest future directions.
Although not explicitly stated, the following hypothesis is strongly implied in the agroforestry literature: Appropriate agroforestry systems improve soil physical properties, maintain soil organic matter, and promote nutrient cycling. It is also frequently mentioned that agroforestry systems require less purchased inputs and can reclaim degraded lands in the tropics. Many traditional agroforestry systems in the tropics involve the use of trees and shrubs on crop and pasture lands (see von Maydell, this volume; Le Houerou, this volume). Farmers often claim that these trees (many of which are nitrogen-fixing species) improve the yields of associated crops (Figure 1).
Notwithstanding the importance of such favoured systems, agroforestry is considered specially applicable to marginal soils with severe physical, chemical or drought constraints. Most of the evidence in support of the soils-agroforestry hypothesis, however, is observational, qualitative or extrapolated from other systems. Many of the quantitative data on the effect of trees on soil properties are based on natural systems or pure plantation forestry (Lundgren, 1978; Chijioke, 1980; Singh, 1982; Sanchez et al, 1985; Vitousek and Sanford, 1986; Andriesse, 1987). Similarly, most of the effects of agronomic practices such as mulching are often taken from crop-production data. Given the critical interaction between trees and crops or pastures, soil-agroforestry relationships should be studied in situations where such interactions take place, and as a function of time. It is the author's proposal that changes in soil properties with time caused by agroforestry systems in well-characterized, long-term data sets be considered as the appropriate test for such hypotheses.
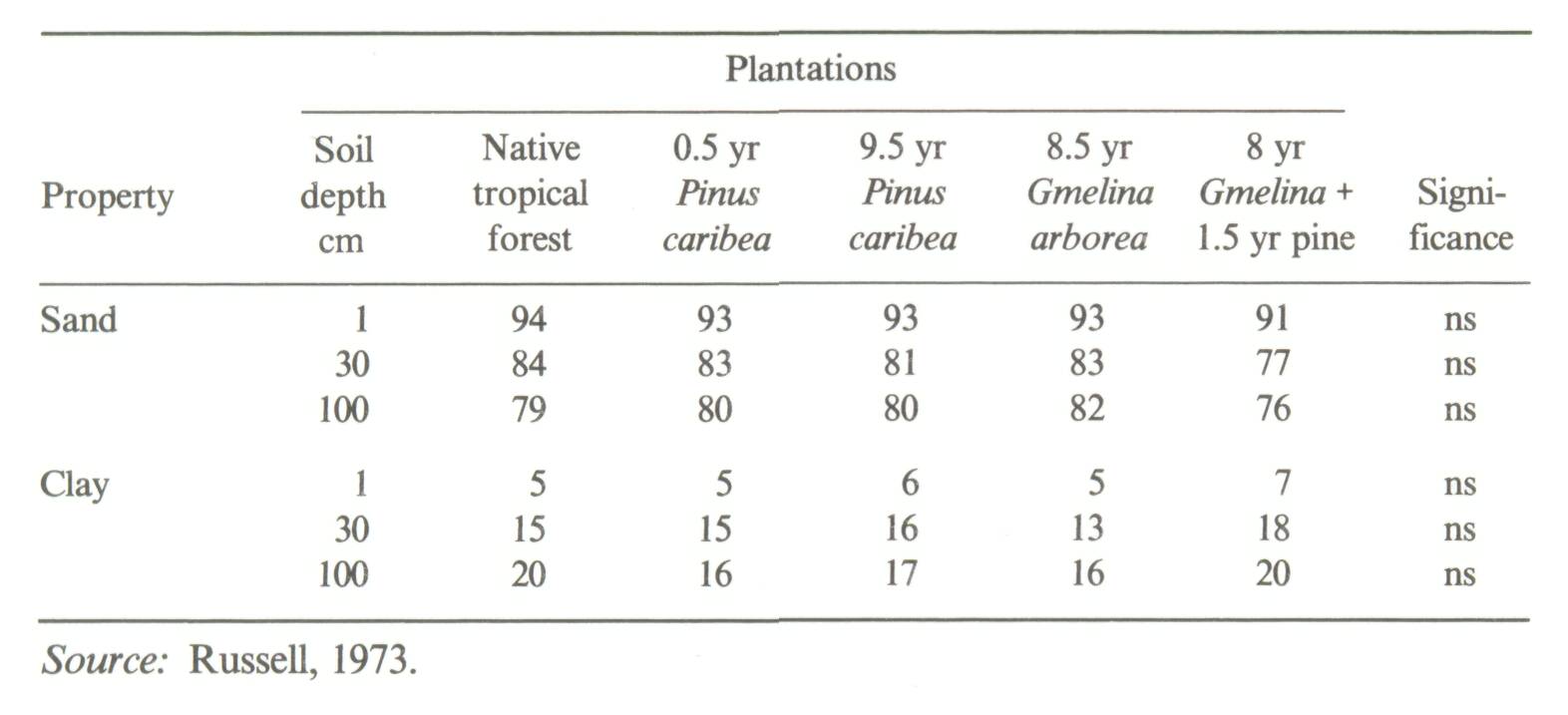
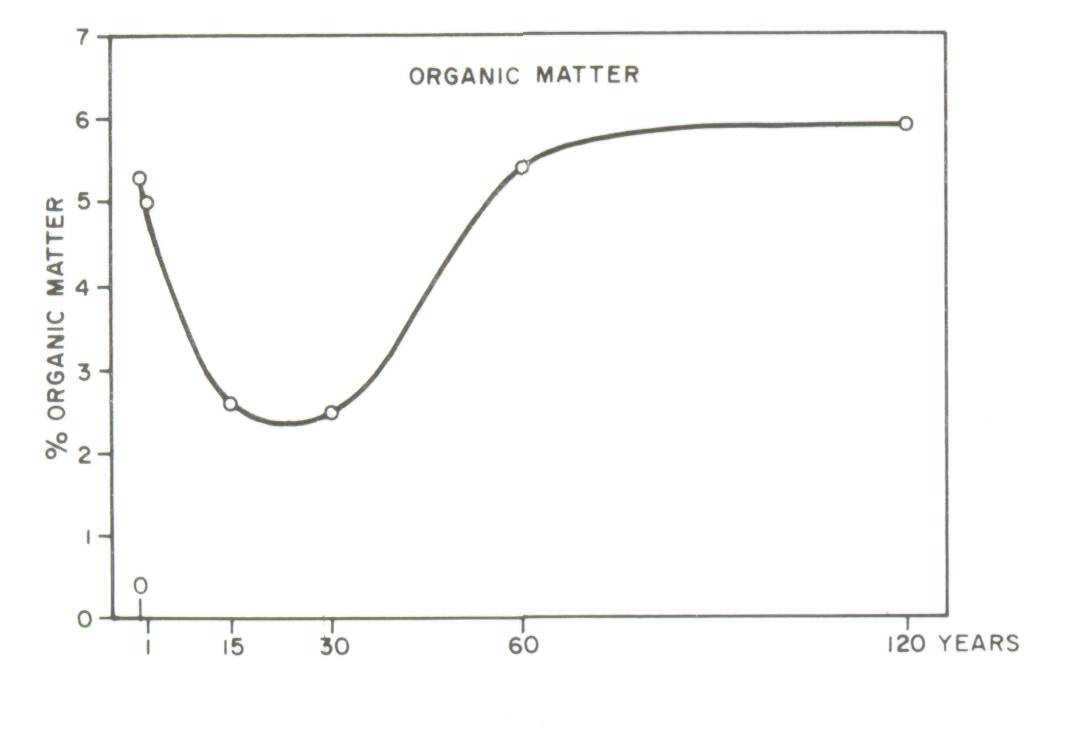
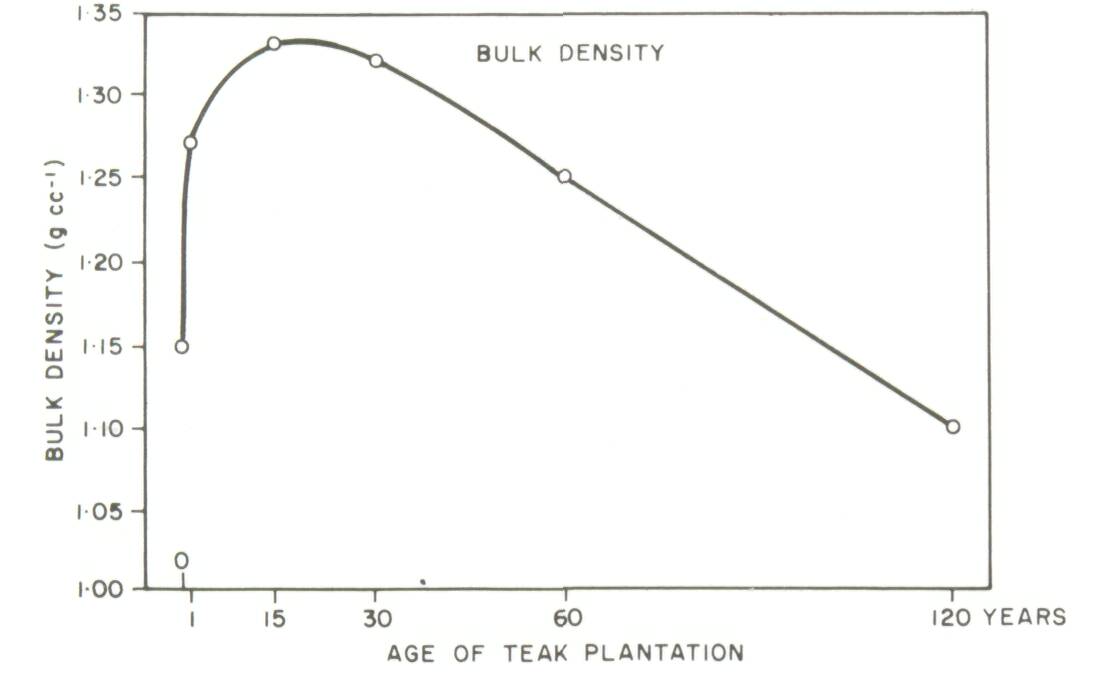
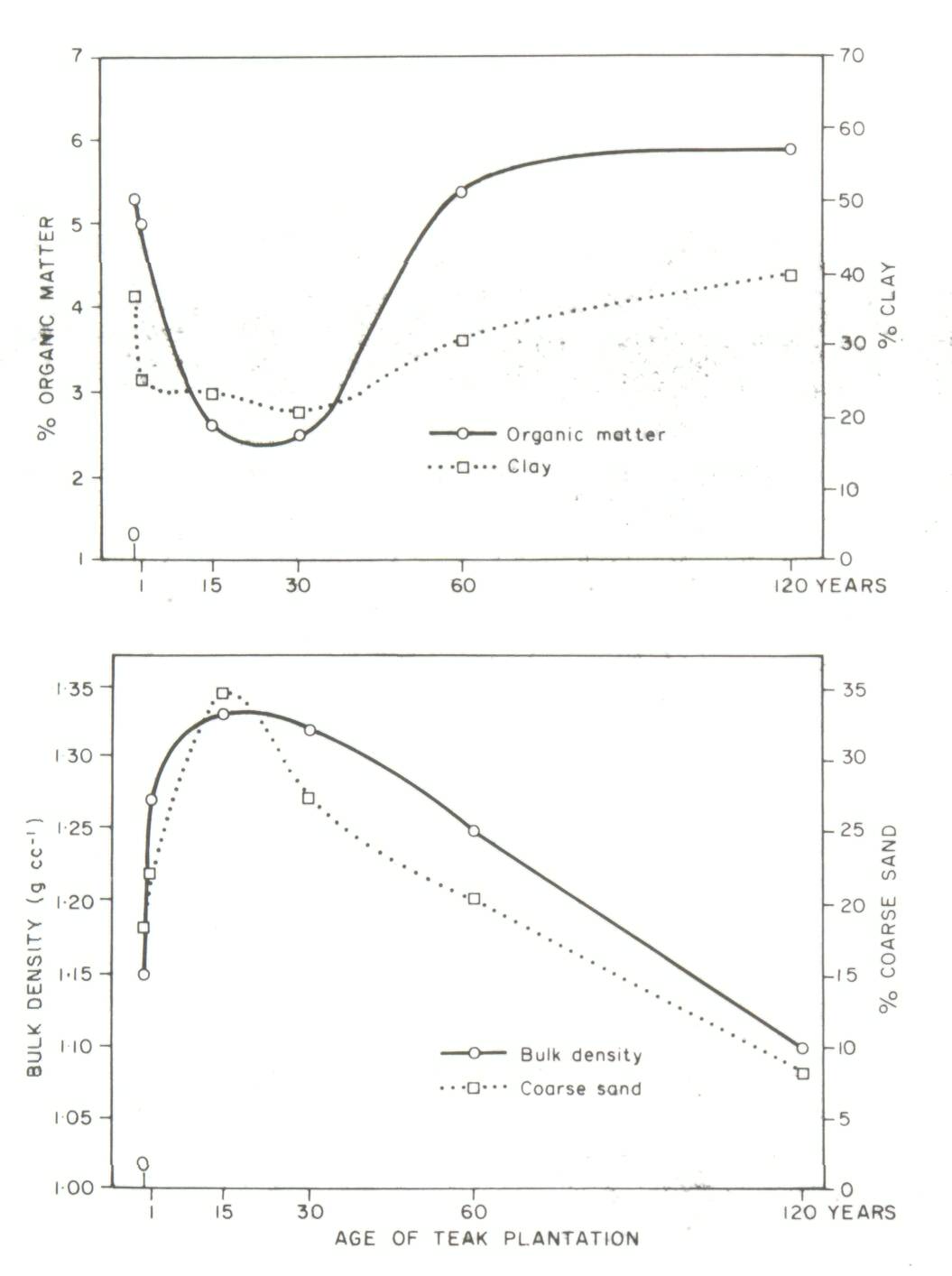
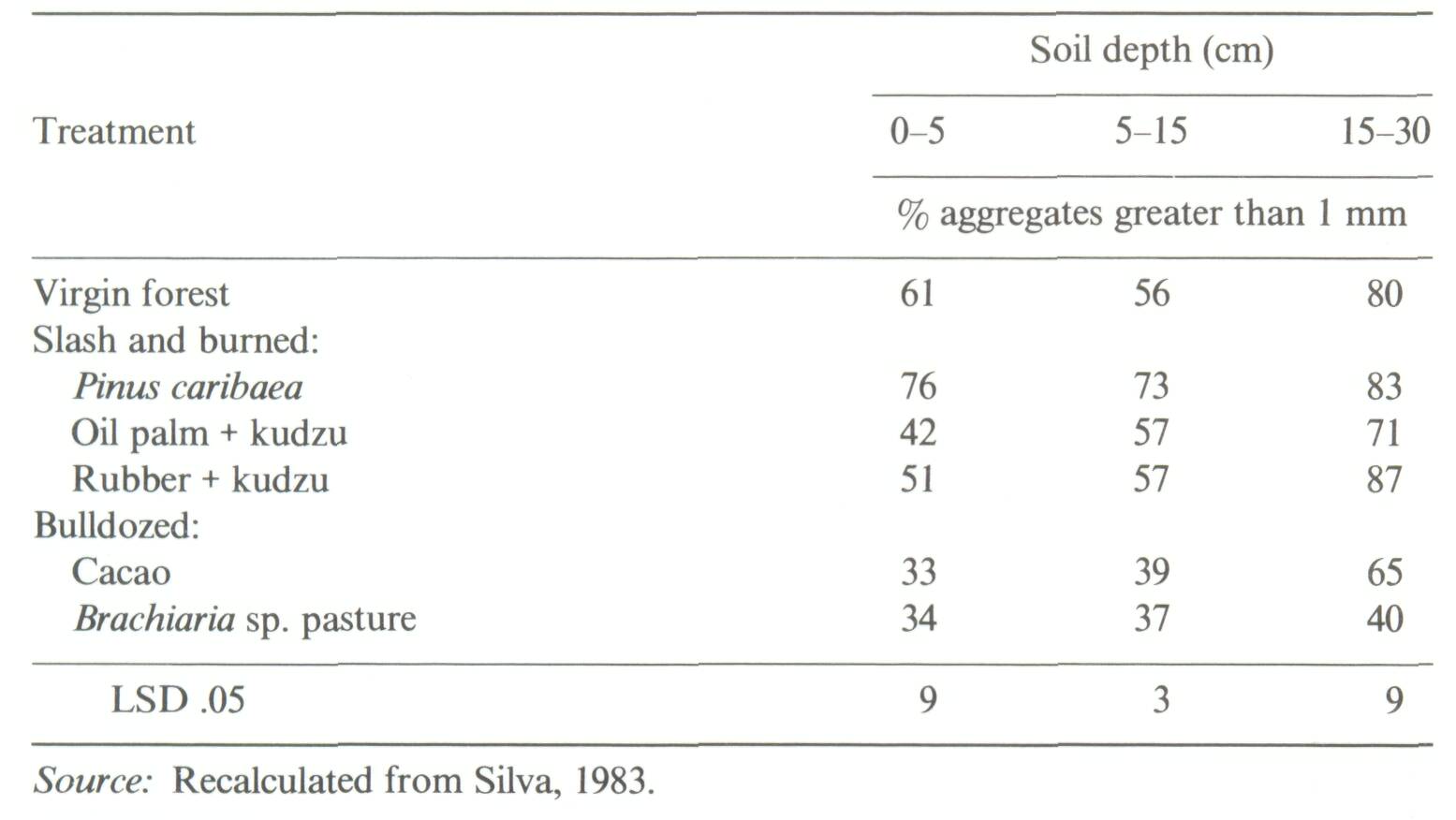
Two types of soil-dynamics data sets were recognized in a recent review (Sanchez et al, 1985): Type I, where changes in soil properties are monitored with time on the same site, and Type II where soils of nearby sites of known dates of planting are sampled at the same time. Well-characterized, replicated Type I experiments are preferred, but they are scarce. The problem with Type II experiments is that the initial conditions and soil properties of the sites are unknown. Thus observations at a later stage may be the result of different initial soil properties or the effect of management systems. The main advantage of a Type II experiment is that it can yield results in a much shorter time as compared to a Type I experiment. Type II experiments can be used to obtain an objective assessment of the effects of an agroforestry system or technology on soil properties, provided comparisons are made on sites with (1) similar soils classified according to soil taxonomy, and (2) little difference in soil texture with depth between sites. In addition, other land factors such as climate, position in landscape and slope aspect should be similar among sites. The greater the variability of a site, the greater will be the need for careful sampling and adequate replication. Particle-size distribution in the profile is a soil property very difficult to change with management or time, and should therefore be a good indicator of whether the soils were similar when the comparisons started. Even in cases of topsoil erosion, particle-size distribution with depth, coupled with the necessary morphological descriptions for classification, can provide reasonable assurance that initial soil properties were the same. An example from the work of Russell (1983) in Ultisols of the Brazilian Amazon is given in Table 1. Differences in sand and clay content between nearby sites were small and not statistically significant. The effects of agroforestry on soil properties can be adequately estimated from such Type II data sets. On the other hand, Type II data sets showing major textural differences between sites, or those providing no soil profile data at all, cannot be used to test the soil-agroforestry hypothesis. An example is a thorough study of teak plantations of up to 120 years of age in Kerala, India. Jose and Koshy (1972) reported major changes in soil organic matter and bulk density with age of teak plantations (Figure 2). Their soil-profile data, however, showed considerable differences in clay and sand content among the various sites supporting plantations of different ages. Clay content is usually correlated with organic matter content, while bulk density is positively related to coarse-sand content. It appears likely that the values for organic matter and bulk density were a function of textural differences rather than the effects of teak plantations over time. Jose and Koshy's data, when drawn in Figure 3, do not support the conclusion that age of teak plantation had a major effect on soil organic matter and bulk density.
An appreciation and application of this fundamental concept of Type I and II experiments will be a major step towards obtaining objective and valid comparisons of the effects of agroforestry systems on soil properties under a variety of environmental and site conditions.
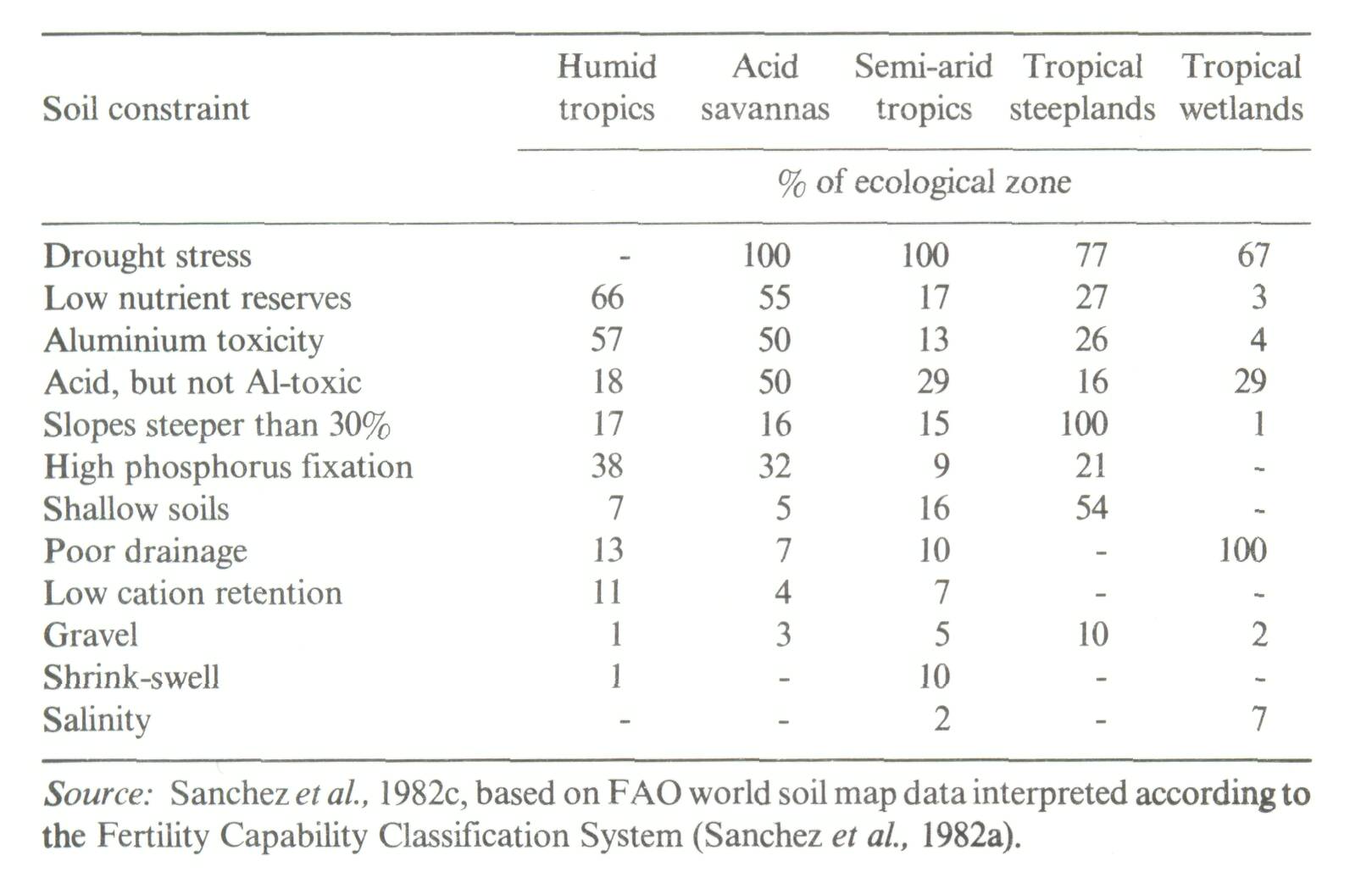
The effects of soils per se on agroforestry systems also deserve more rigorous scrutiny. Major advances have been made on the properties, distribution and constraints of soils in the tropics during the last decade. The widespread use of a quantitative soil taxonomy system (Soil Survey Staff, 1975), the completion of the FAO world soil map, and many local soil surveys provide a vastly improved data base on soil properties and their geographical distribution. Some widely held views of the 1960s are now contradicted by quantitative knowledge. For example, generalizations such as "most tropical soils are low in organic matter", or that such soils "turn into laterite after clearing" are not supported by the data. Soil maps or relatively complicated taxonomic names often convey little to agronomists or foresters. Technical classification systems are used to interpret soil taxonomy information in terms relevant to specific users. Foresters frequently use the Site Index, which is a technical land classification system based on the observed rate of tree growth. The Fertility Capability Classification (FCC) system is a technical system that provides interpretations in terms of soil constraints to plant growth (Buol et al, 1975; Sanchez et al, 1982a). An example of the geographical extent of soil constraints by major agro-ecological zones is shown in Table 2, where the relative importance of each FCC constraint varies between the humid tropics, acid savannas, semi-arid tropics, steeplands and wetland regions. Local knowledge of soil taxonomy and FCC constraints, however, is more relevant than such world-wide figures. Local data on soil constraints can be included in the diagnosis, design or improvement of agroforestry systems. Similarly, the interpretation of agroforestry data without regard to the soils where the research was conducted severely limits its potential applicability.
The three components of the soil-agroforestry hypothesis are now examined using data sets that meet the Type I or acceptable Type II experiment criteria and are related to soil properties and constraints. Examples from soils of the humid, subhumid and semi-arid tropics are used.
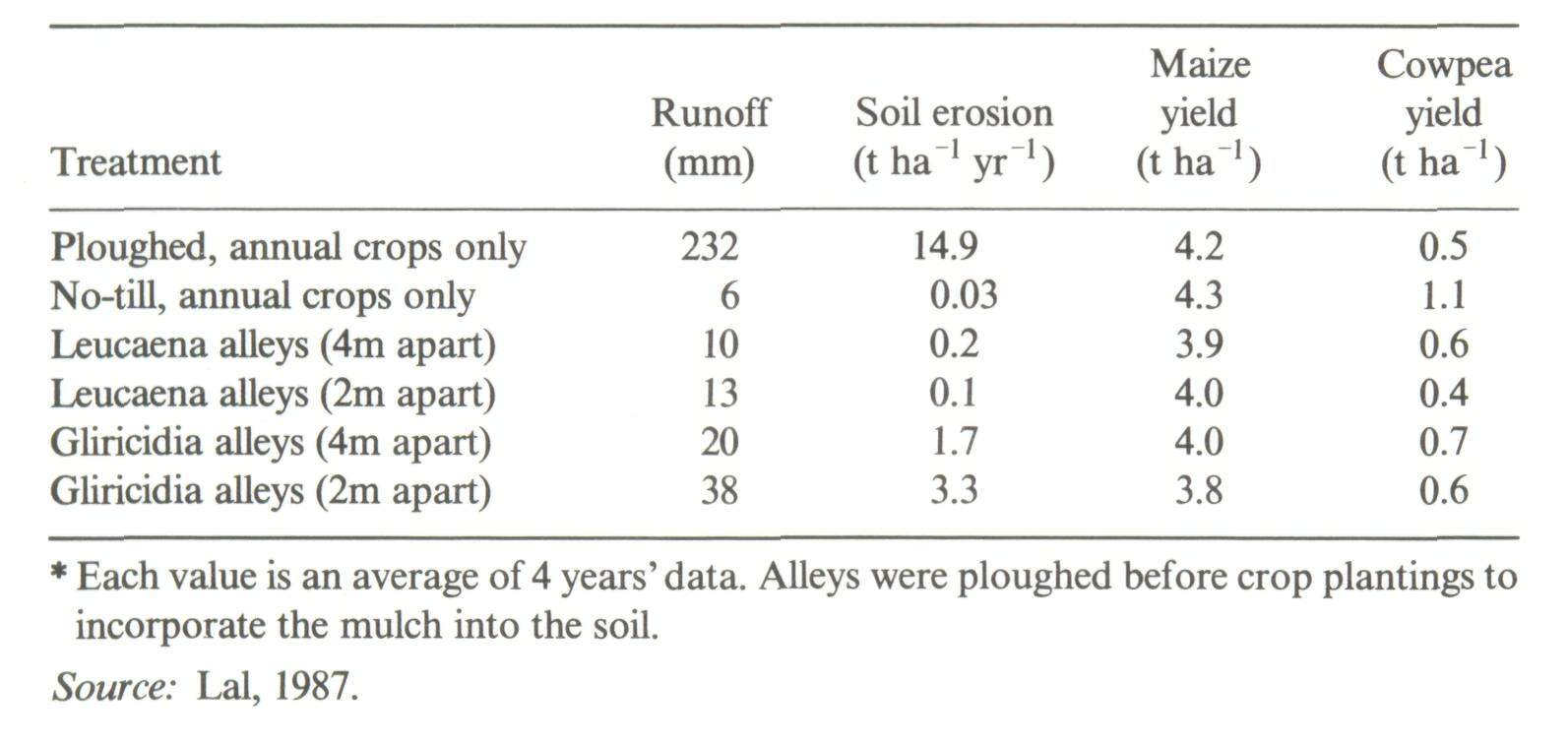
The fundamental reason why agroforestry systems are perceived to improve soil properties is the protection a tree cover gives the soil against surface compaction, runoff and erosion (Young, 1986a). Agroforestry systems can contain one or more such covers (also referred to as canopies): (1) a tree-top canopy, (2) aground cover provided by annual crops or pasture, and (3) a surface-litter layer produced by the vegetation when any or all of its components are fully established. The onset, duration and thickness of ground cover of each type varies widely in agroforestry systems. Two ICRAF reviews reported ample data showing that the presence of trees decreases soil erosion but no direct evidence from agroforestry systems (Nair, 1984; Young, 1986c). Most of the information is based on what happens when forests are cut down, but not on what happens when trees are planted in combination with crops or pastures. Although it is generally believed that a plant cover is the best way to protect the soil from erosion, it need not always be so (Wiersum, 1984). The question is, to what extent do individual agroforestry systems produce plant covers that can reduce erosion, how soon, and for how long a period of time? Most forested land in the humid and subhumid tropics that has been cleared by mechanical means suffers some physical deterioration (Seubert et al., 1977; Lal et al, 1986) ranging from outright topsoil removal to mild surface-soil compaction. The detrimental effects of improper land clearing similarly affect crop, pasture, or tree growth (Sanchez et al, 1985). The land-clearing method used, therefore, is a major starting point for interpreting changes in soil physical properties with time. Furthermore, improvement in physical properties implies that soils need physical improvement. This is less the case with well-aggregated Oxisols and Andepts and in well-aggregated oxic Ultisols and Alfisols than with many sandy Alfisols and Ultisols that have very unstable structural aggregates. One fact not often mentioned in the agroforestry literature is that most trees develop a soil-protecting plant canopy more slowly than annual crops or pastures (Broughton, 1979). Most humid tropical tree crops require at least two years to close their canopy, whereas annual crops provide adequate cover within 30 to 45 days and pastures within two to six months. Although there is sufficient evidence that fully developed tree canopies reduce runoff and erosion losses to levels similar to the original forest vegetation, the soil is particularly exposed during the tree-establishment phase unless an annual crop or a legume ground cover is planted shortly after land clearing. A Type II study of a five-year-old Acacia auriculiformis plantation under a lowland humid climate in Java compared the effects of tree&canopy removal, undergrowth and litter on soil erosion. The tree-canopy itself had little effect on soil erosion and the effect of undergrowth was small. Litter cover alone reduced soil erosion by 95 percent as compared with bare soil (Wiersum, 1985). To maintain continuous litter, however, fresh inputs from the tree/shrub canopies were required. There is ample information on the effects of annual crop canopies on runoff and erosion from a variety of soils and slopes (Roose, 1970; Lal, 1975). Agroforestry systems may provide adequate soil protection at the critical establishment phase, but mainly by crops or legume ground covers not by the trees. This will be an important factor in the design of agroforestry technologies for minimizing soil erosion. Once established, however, an agroforestry technology can provide significant soil protection. This is shown in a successful alley-cropping trial on a pH 6 sandy Entisol at Ibadan, Nigeria which is highly susceptible to soil erosion (Lal, 1987) (Table 3). The existence of trees did not affect maize yields, but did reduce cowpea yields considerably as compared with the best management for annual crop rotations on the site.
In the semi-arid tropics, Acacia albida has a reputation for being a soil improver, as evidenced primarily by increasing crop yields under its deciduous canopy. A review of the literature by Felker (1978) indicates that the soil's water-holding capacity increases under Acacia albida in comparison with nearby sites devoid of such trees. The reports by Charreau and Vidal (1965), Dancette and Poulain (1969) and Dugain (1960) do not supply detailed soil characterization from the different sites, although it may be reasonable to assume similarity. The effect of increased water-holding capacity is believed to be a consequence of higher topsoil organic-matter content under Acacia albida on very sandy
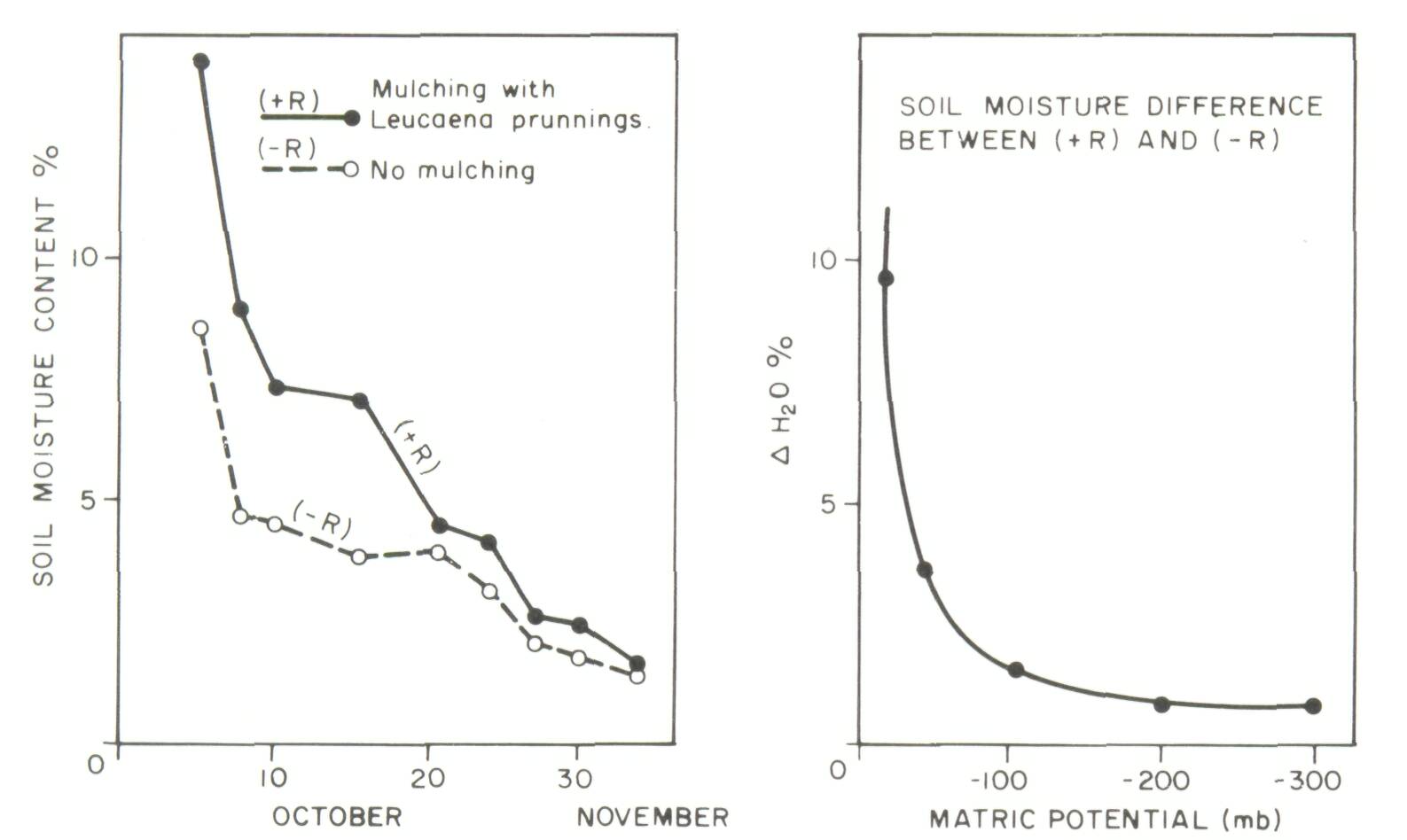
Alfisols which are very low in organic matter in the natural state. It is interesting to note that Poschen (1986) concluded that on poorly drained sites, enhanced crop production under Acacia albida canopies was due to improved drainage. No Type I experiment on the effect of Acacia albida on soil properties was reported in Felker's review. In addition, the lack of adequate descriptions and/ or classifications of soils and the absence of acceptable Type II studies severely limits the validity of many of the inferences about the effects of Acacia albida on soils. Clearly, the widespread generalization that Acacia albida is found mainly in sandy, infertile sites is not true. Miehe (1986) describes Acacia albida systems on high base status soils in the Sudan. Geostatistical techniques could be tried on fields with randomly occurring Acacia albida trees in the Sahel to separate the observed positive effects of trees on crops growing under them from random soil spatial variability. Type II studies on Acacia albida in the Sahel (Jung, 1966) and Prosopis cineraria in Rajasthan, India (Mann and Saxena, 1980) have revealed increased clay content beneath the canopies of these trees. Such observations could be due to the trees acting as a physical barrier and trapping clay particles during the many dust storms that are prevalent in these regions. It is also possible that the tree's presence on a particular site may be due to the higher clay content on the site to begin with. In either case, such observations provide a good basis for well-designed Type I or acceptable Type II experiments to pinpoint the factors operating under various site and environmental conditions. Alley cropping, growing leguminous shrubs as a source of mulch within rows along crop fields (see Kang and Wilson, this volume), is one of the most publicized agroforestry options for the humid and subhumid tropics. In spite of the attention presently given to alley cropping in the tropics, published information relating the effect of alley cropping to improved soil physical properties is scarce. Kang et al. (1985) reported that the additions of Leucaena prunings in a maize alley-cropping system substantially increased moisture retention of the topsoil (Figure 4). The improvement is quite important because the soil in question is a sandy gravelly Entisol which is very credible and subject to high temperature fluctuations. Table 3 provides data on the effect of alley cropping on soil erosion.
Furthermore, the protection of the soil surface by mulch layers, whether a layer from free-growing trees or prunings from alley cropping, is a dynamic process, affected by the quantity, timing and rate of decomposition of mulch inputs. Very little quantitative work on these aspects has been done, although their importance is acknowledged in the literature on natural systems (Swift et al., 1979). In agroforestry systems, the three factors (quantity, timing and decomposition rate) may determine whether a large initial addition of mulch will smother weed growth while permitting normal crop development. Rapidly decomposing mulch may not stay on the surface for a sufficiently long period, and thus may provide only temporary cover. The rate of decomposition of organic materials, often called "litter quality", depends on several factors, among which the lignin/nitrogen ratio and total soluble polyphenolic content of leaf blades are believed to be important (Melillo et al, 1982; Spain and Le Feuvre, 1987). No effects of mulch litter quality on agroforestry systems have been found in the available literature in Type I or appropriate Type II experiments. An assessment of parameters that would predict the time required for a sufficiently thick mulch layer to provide adequate soil cover needs to be undertaken in promising agroforestry systems using properly designed soil-dynamics experiments.
Agroforestry systems are believed to increase, or at least maintain, the organic-matter levels of the soil (Young, 1986a,b; Young et al., 1986). The range in soil organic matter (SOM) content in the tropics is similar to that found in temperate regions (Sanchez and Buol, 1975; Sanchez et al., 1982b) and the equilibrium content of any soil is a function of organic inputs (addition) and decomposition rates (Greenland and Nye, 1959). Topsoil organic carbon levels usually decrease after clearing tropical forest and reach a new equilibrium after several years of different levels of organic inputs and decomposition rates. Organic-matter dynamics have been reported on annual crops (Sanchez et al., 1983), pastures (Serrao et al., 1979), and tree crops (Sanchez et al, 1985) for various tropical ecosystems. The known beneficial effects of SOM on crop production were summarized by various workers, including Allison (1973) and Swift and Sanchez (1984). In short, SOM is (1) a source of inorganic nutrients for plants, (2) a substrate for micro-organisms, (3) an ion-exchange material, (4) a factor in soil aggregation and root development, and consequently (5) a factor in soil and water conservation. From a practical standpoint, it is desirable to have a nutrient source that synchronizes nutrient release with plant-growth demands. This is the basis of our chemical-fertilizer technology. According to Swift (1984, 1985), successful management of organic inputs should be directed at a nutrient-release pattern in synchrony with the crop's nutrient uptake pattern, taking into consideration the soil's ability to regulate related processes such as leaching and denitrification. This "SYNCH" concept is one of the fundamental tenets of the world-wide Tropical Soils Biology and Fertility (TSBF) programme (Swift, 1984; Swift and Sanchez, 1984). Holland and Coleman (1987) reported slower decomposition of surface litter or mulch in no-till than in tilled systems due to the presence of a higher proportion of fungi in the mulch layer as compared to the decomposer community present in the soil. Increased fungal decomposition may enhance organic-matter retention because fungi retain a higher proportion of metabolized carbon than bacteria (Adu and Oades, 1978). Also, decomposition by fungi can produce more recalcitrant organic fractions than bacteria (Mayauden and Simonart, 1963). Current work on SOM is focusing on functional pools as opposed to total SOM levels (Parton et al., 1983; Sanchez and Miller, 1987). Consideration of these fractions, defined on the basis of their turnover rates in the soil, may have more relevance for evaluating agroforestry technologies than looking at total soil organic matter content. Parton et al. (1983) define three functional pools: active SOM (turnover rate 0-3 years), slow SOM (turnover up to 25 years) and passive SOM (about 1,000 years). The underlying hypothesis is that maximization of the active and slow SOM pools will result in greater "activity" of SOM with regard to nutrient release, complexing Al, and soil structure. In soil testing for fertilizer use, we no longer analyse "the bones" but "the bloodstream" (for example, not total P, but available P). A similar goal for organic matter is now being set by soil scientists involved in TSBF.
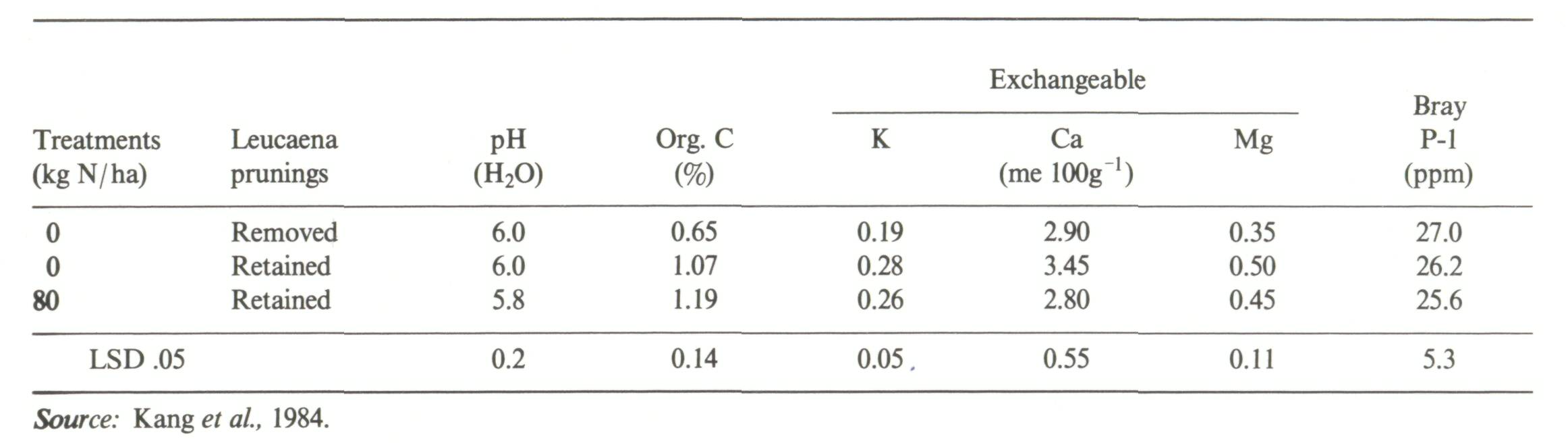
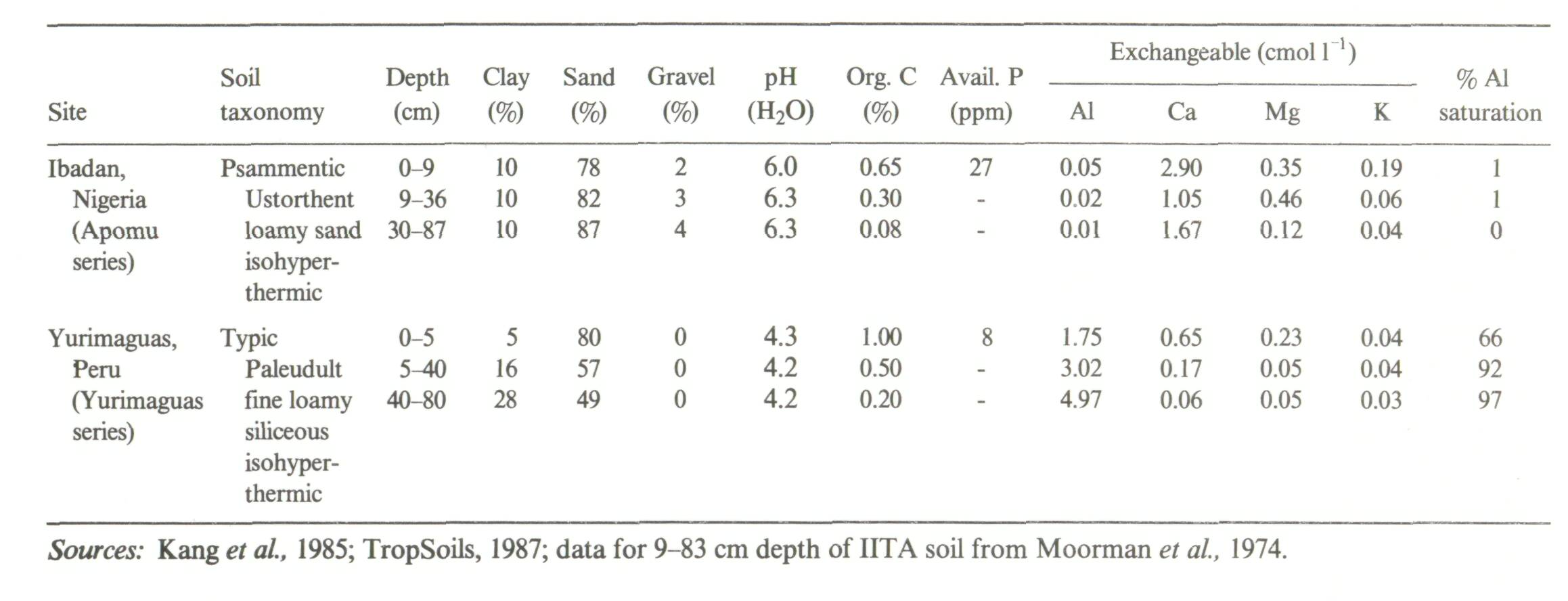
One of the advantages commonly attributed to agroforestry technologies is the potential for soil fertility improvement via more efficient cycling of nutrients (Nair, 1984), and it is often recommended to include nitrogen-fixing trees and shrubs in such technologies (Nair et al, 1984; Lundgren and Nair, 1985; Young, 1987). Good evidence exists from Type II experiments for the nutrient-cycling potential of agroforestry systems on Alfisols and Andepts of moderate to high fertility. Such systems as the Erythrina poeppigiana shade trees over Coffea arabica in Costa Rica are a good example (Glover and Beer, 1986; Russo and Budowski, 1986; Alpizar et al, 1986). Similarly, Roskoski (1981) reported that Inga jinicuil, a leguminous shade tree in coffee plantations, fixed around 40 kg N ha-1 yr-1. Increases in nodule biomass and function appeared to be positively correlated with increasing P-fertilizer applications to the coffee, suggesting that phosphorus deficiencies had to be ameliorated prior to adequate functioning of nodules. Juo and Lal (1977) compared the effects of a leucaena fallow versus a bush fallow on selected soil chemical properties on an Alfisol in western Nigeria in a Type I experiment. After three years, during which leucaena was cut annually and left as mulch, the leucaena fallow resulted in significantly higher effective cation exchange capacity and levels of exchangeable Ca and K as compared to the bush fallow. Similarly, in a Type II experiment, an agroforestry system involving oil palm with leguminous cover crops (Centrosema pubescens and Pueraria phaseoloides) appeared more efficient at nutrient cycling than an oil-palm plantation with no cover crop. In addition to fixing about 150 kg N ha-1 yr-1, the loss of nitrate nitrogen via leaching was significantly lower in the agroforestry system than in the sole crop of oil palm (Agamuthu and Broughton, 1985). After six years of alley cropping with Leucaena leucocephala on a pH 6 Entisol, plots receiving prunings had a higher nutrient status and twice the organic-matter content than plots not receiving prunings (Kang et al, 1984) (Table 5). This is a highly successful agroforestry system. Attempts to extrapolate the IITA-leucaena alley cropping model to a highly weathered, sandy Ultisol in the Amazon basin of Peru, however, were not as successful (TropSoils, 1987). Table 6 shows a comparison of some of the soil properties between the Nigerian and Peruvian sites. The main differences are (1) the susceptibility of Leucaena leucocephala to Al toxicity, and (2) the low nutrient base content of the Yurimaguas subsoil which reduced the recycling pool. The poor performance of leucaena in acid soils had been anticipated and a number of other woody species were evaluated in alley-cropping trials. These include Inga edulis, Codariocaly (Desmodium) gyroides and Erythrina spp. Although yield data and soil properties are still under analysis, Inga edulis appears to be a highly promising agroforestry species for acid soils in the tropics (Figure 5). Although alley cropping works well in moderately fertile soils, current experience suggests that it will be necessary to use inputs such as lime, and possibly P, to allow successful establishment of alley-cropping species and subsequent recycling of nutrients on acid infertile Ultisols and Oxisols (TropSoils, 1986). Much further study is required before this particular agroforestry system can be considered widely applicable to the humid and subhumid tropics. Some tree and shrub species can selectively accumulate certain nutrients even in soils containing very low amounts of these nutrients. Thus palms and palm litter are rich in potassium (Folster et al., 1976), tree ferns accumulate nitrogen (Mueller-Dombois et al, 1984), Cecropia spp. appeared to accumulate Ca and P on acid sites (Odum and Pigeon, 1970) and Gmelina arborea accumulates Ca (Sanchez et al, 1985). It is important for researchers looking to incorporate such nutrient-conserving species into agroforestry technologies to appreciate the fact that such plant-nutrient responses will change with location and site/soil characteristics (Golley, 1986).
Agroforestry as a major approach to sustainable land use in the tropics is now a widely acknowledged concept. Many of its positive attributes relate to the management and conservation of marginal soils of the tropics. While evidence exists for the beneficial effects on soils of certain agroforestry technologies (especially on more fertile soils), there is a tendency for over-generalization and extrapolation of soil productivity and sustainability benefits of agroforestry systems to other more marginal sites. The time has come to bring science into the picture and systematically test the effects of agroforestry systems on different soils, and vice versa. A soil-dynamics methodology is proposed as a framework for such testing. The above-and below-ground interactions between trees and crops or pastures are likely to provide different results from those obtained in forests, cropped fields or pastures. Recognition of what the major soil constraints are in specific areas would improve the design of agroforestry systems. Science-based soil-agroforestry research will provide a realistic site-specific appraisal of whether agroforestry systems improve soil physical properties, maintain soil organic matter or promote nutrient cycling.
Paper No. 11163 of the Journal Series of the North Carolina Agricultural Research Service, Raleigh, N.C., U.S.A. The author is deeply indebted to Mr Erick CM. Fernandes, graduate student, for assistance in compiling materials for the paper.
Adu, J.K. and J.M. Oades. 1978. Utilization of organic materials in soil aggregates by bacteria and fungi. Soil Biology and Biochemistry 10:117-122. Agamuthu, P. and W.J. Broughton. 1985. Nutrient cycling within the developing oil palm-legume ecosystem. Agriculture, Ecosystems and Environment 13:111-123. Allison, F.E. 1973. Organic matter and its role in crop production. Amsterdam: Elsevier. Alpizar, L., H.W. Fassbender, J. Heuveldop, H. Folster and G. Enriquez. 1986. Modelling agroforestry systems of cacao (Theobroma cacao) with laurel (Cordia alliodora) and poro (Erythrina poeppigiana) in Costa Rica. Inventory of organic matter and nutrients. Agroforestry Systems 4: 175-189. Andriesse, J. P. 1987. Monitoring project of nutrient cycling soils used for shifting cultivation under various climatic conditions in Asia. Final Report. Joint KIT/EEC Project No. TSD-A- 116-NL. Royal Tropical Institute, Amsterdam, The Netherlands. Broughton, W.J. 1979. Effects of various covers on soil fertility under Hevea brasiliensis and on growth of the tree. Agro-Ecosystems 3: 147-170. Buol, S.W., P. A. Sanchez, R.B. Cate and M.A. Granger. 1975. Soil fertility capability classification. In E. Bornemisza and A. Alvarado (eds.), Soil management in tropical America. Raleigh, NC: North Carolina State University. Charreau, C. and P. Vidal. 1965. Influence de I'Acacia albida Del. sur le sol, nutrition minerale et rendements des mils Pennisetum au S6negal. Agron. Trap. 6-7: 626-660. Chijioke, E.0.1980. Impact on soils of fast-growing species in lowland humid tropics. FAO Rome:FAO Forestry Paper 21. Dancette, C. and J.F. Poulain. 1969. Influence of Acacia albida on pedoclimatic factors and crop yields. African Soils 14: 143-184. Dugain, G. 1960. Rapport de Mission au Niger. Centre de Peclologie de HANN - Dukan. (Mimeo.) Felker, P. 1978. State of the art: Acacia albida as a complementary permanent intercrop with annual crops. University of California, Riverside. Fernandes, E.C.M., A. OTctingati and J. Maghembe. 1984. The Chagga homegardens: a multistoried agroforestry cropping system on Mt. Kilimanjaro, Tanzania. Agroforestry Systems 2: 73-86. Folster, H., G. de las Salas and P. Khanna. 1976. A tropical evergreen forest site with perched water table. Magdalena Valley, Colombia. Biomass and bioelement inventory of primary and secondary vegetation. Oecol. Plant. 11: 297-320. Glover, N. and J. Beer. 1986. Nutrient cycling in two traditional Central American agroforestry systems. Agroforestry Systems 4: 77-87. Golley, F.B. 1986. Chemical plant-soil relationships in tropical forests. J. Tropical Ecol. 2:219-229. Greenland, D.J. and P.H. Nye. 1959. Increases in the carbon and nitrogen contents of tropical soils under natural fallows. Soil Sci. 10: 284-299. Holland, E.A. and D.C. Coleman. 1987. Litter placement effects on microbial and organic matter dynamics in an agroecosystem. Ecology 68: 425-433. Jose, A.I. and M.M. Koshy. 1972. A study of the morphological, physical and chemical characteristics of soils as influenced by teak vegetation. Indian Forester 98: 338-348. Jung, G. 1966. Etude de Tinfluence de VAcacia albida (Del.) sur les processus microbiologiques dans le sol et sur leurs variations saisonnieres. Centre ORSTOM-Dakar Senegal. (Mimeo.) Juo, A.S.R. and R. Lal. 1977. The effect of fallow and continuous cultivation on the chemical and physical properties of an Alfisol in Western Nigeria. Plant Soil 47: 567-584. Kang, B.T., G.F. Wilson and T.L. Lawson. 1984. Alley cropping: A stable alternative to shifting cultivation. Ibadan, Nigeria: IITA. Kang, B.T., H. Grimme and T.L. Lawson. 1985. Alley cropping sequentially cropped maize and cowpea with Leucaena on a sandy soil in Southern Nigeria. Plant Soil 85: 267-277. Kellman, M. 1979. Soil enrichment by neotropical savanna trees. J. Ecol. 67: 565-577. Lal, R. 1975. Role of mulching techniques in soil and water management. IITA Technical Bulletin No. 1, Ibadan, Nigeria. .1987. Managing the soils of sub-Saharan Africa. Science 236: 1069-1076. Lal, R., P. A. Sanchez and R.W. Cummings, Jr. (eds.). 1986. Land clearing and development in the tropics. Boston: A.A. Balkema. Lundgren, B. 1978. Soil conditions and nutrient cycling under natural and plantation forests in Tanzanian highlands. Report in Forest Ecology and Forest Soils No. 31, Swedish University of Agricultural Science, Uppsala. Lundgren, B. and P.K.R. Nair. 1985. Agroforestry for soil conservation. In S.A. El-Swaify, W.C. Moldenhauer and A. Lo (eds.), Soil erosion and conservation. Ankeny, Iowa: Soil Conservation Society of North America. Mann, H.S. and S.K. Saxena (eds.). 1980. Khejri (Prosopis cineraria) in the Indian Desert. CAZRI Monograph No. 11. Jodhpur: Central Arid Zone Research Institute. Mayauden, J. and P. Simonart. 1963. Humification des microorganismes marques par I4C dans le sol. Annales de I'lnstitut Pasteur 105: 257-266. Michon, G., F. Mary and J. Bompand. 1986. Multistoried agroforestry garden system in West Sumatra, Indonesia. Agroforestry Systems 4: 315-338. Miehe, S. 1986. Acacia albida and other multipurpose trees on the Fur farmlands in the Jebel Maia highlands, Western Dafur, Sudan. Agroforestry Systems 4: 89-119. Melillo, J.M., J.D. Aber and J.F. Muratore. 1982. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecol. 63: 621-626 Mongi, H.O. and P.A. Huxley (eds.). 1979. Soils research in agroforestry. Nairobi: ICRAF. Moormann, F.R., R. Lal and A.S.R. Juo. 1975. The soils of IITA. Technical Bulletin No.3. IITA, Ibadan, Nigeria. Mueller-Dombois, D., P.M. Vitousek and K.W. Bridger. 1984. Canopy dieback and ecosystem processes in Pacific forests. Hawaii Botanical Science Paper 44, University of Hawaii,Manoa. Nair, P.K.R. 1984. Soil productivity aspects of agroforestry. Nairobi: ICRAF. .1987. Soil productivity under agroforestry. In H. Gholz (ed.). Agroforestry: realities, possibilities and potentials. Dordrecht, Netherlands: Martinus Nijhoff (in press). Nair, P.K.R., E.C.M. Fernandes and P. Wambugu. 1984. Multipurpose trees and shrubs for agroforestry. Agroforestry Systems 2: 145-163. Odum, H.T. and R.F. Pigeon (eds.). 1970. A tropical rainforest. Vol. III. Washington, D.C.: Office of Information Services. U.S. Atomic Energy Commission. Parton, W.J., D.W. Anderson, C.V. Cole and J.W.B. Stewart. 1983. Simulation of soil organic formations and mineralization in semiarid agroecosystems. In R.R. Lowrance, R.L. Todd, L.E. Asmussen and R.A. Leonard (eds.), Nutrient cycling in agricultural ecosystems. College of Agriculture Experimental Station, Special Publication No. 23, University of Georgia, Athens. Poschen, P. 1986. An evaluation of the Acacia albida-based agroforestry practices in the Hararghe highlands of Eastern Ethiopia. Agroforestry Systems 4: 129-143. Riha, S.J., B.R. James, G.P. Senesacand E. Pallant. 1986. Spatial variability of soil pH and organmatter in forest plantations. Soil Sci. Soc. Am. J. 50: 1347-1352. Roose, E. 1970. Importance relative de l'erosion, du drainage oblique et vertical dans la pédogénése d'un sol ferralitique de moyenne Cote d'Ivoire. Cah. ORSTOM Ser. Pédol. 8: 469-482. Roskoski, J.P. 1981. Modulation and nitrogen fixation by Inga jinicuil, a woody legume in coffeeplantations. I. Measurements of nodule biomass and field acetylene reduction rates. Plant Soil 59: 201-206. Russel, C.E. 1983. Nutrient cycling and productivity in native and plantation forests in Jari Florestal, Para, Brasil. Ph.D. thesis, Institute of Ecology, University of Georgia, Athens. Russo, R.O. and G. Budowski. 1986. Effect of pollarding frequency on biomass of Erythrina poeppigiana as a coffee shade tree. Agroforestry Systems 4: 145-162. Sanchez, P.A. 1979. Soil fertility and conservation considerations for agroforestry systems in the humid tropics of Latin America. In H.O. Mongi and P.A. Huxley (eds.), Soils research in agroforestry. Nairobi: ICRAF. Sanchez, P.A. and S.W. Buol. 1975. Soils of the tropics and the world food crisis. Science 188:598-603. Sanchez, P.A., W. Couto and S. W. Buol. 1982a. The fertility capability soil classification system: interpretation, applicability and modification. Geoderma 27: 283-309. Sanchez, P. A., M.P. Gichuru and L.B. Gatz. 1982b. Organic matter in major soils of the tropical and temperate regions. Transactions of the Twelfth International Congress of Soil Science, New Delhi 1:99-114. Sanchez, P.A. and R.H. Miller. 1987. Organic matter and soil fertility management in acid soils of the tropics. Transactions of the Thirteenth International Congress of Soil Science, Ham(in press). Sanchez, P.A., J.J. Nicholaides and W. Couto. 1982c. Physical and chemical constraints to food production in the tropics. In G. Bixler, and L. W. Shemilt (eds.), Chemistry and world food supplies: the new frontiers. CHEMRAWN II. Perspectives and recommendations. Los Banos, Philippines: IRRI. Sanchez, P. A., C. A. Palm, C.B. Davey, L.T. Szott and C. E. Russell. 1985. Trees as soil improvers in the humid tropics? In M.G.R. Cannell and J.E. Jackson (eds.), Attributes of trees as crop plants. Huntingdon, U.K.: Institute of Terrestrial Ecology. Sanchez, P. A., J.H. Villachica and D.E. Bandy. 1983. Soil fertility dynamics after clearing a tropical rainforest in Peru. Soil Sci. Soc. Am. J. 47: 1171-1178. Serrao, E. A.S., I.C. Falesi, J.R. Veiga and J. F. Texeira. 1979. Productivity of cultivated pastures in fertility soils of the Amazon of Brazil. In P.A. Sanchez and L.E. Tergas (eds.), Pasture production in acid soils of the tropics. Cali, Colombia: CIAT. Seubert, C.E., P.A. Sanchez and C. Valverde. 1977. Effects of land clearing methods on soil properties and crop performance in an Ultisol of the Amazon jungle of Peru. Trap. Agric. (Trin.) 54: 307-321. Silva, L.F. 1983. Influencia de cultivos e sistemas de manejo nas modificaoes edaficas dos oxisols de tabuleiro (Haplorthox) do sul da Bahia. Belem, Brazil: CEPLAC, Departamento Especial da Amazonia. Singh, B. 1982. Nutrient content of standing crop and biological cycling in Pinuspatula ecosystems. For. Ecol. Management 4: 317-322. Soil Survey Staff. 1975. Soil taxonomy (Agricultural Handbook No. 436). Washington, D.C.:United States Department of Agriculture. Spain, A.V. and R.P. Le Feuvre. 1987. Breakdown of litters of contrasting quality in a tropical Australian rainforest. Journal of Applied Ecology 24: 279-288. Swift, M.J. (ed.). 1984. Soil biological processes and tropical soil fertility: A proposal for a collaborative program of research. Biology International Special Issue 5. Paris:International Union of Biological Sciences. .1985. Tropical soil biology and fertility (TSBF): Planning for research. Biology International Special Issue 9. Paris: International Union of Biological Sciences. Swift, M. J., O. W. Heal and J.M. Anderson. 1979. Decomposition in terrestrial ecosystems. Oxford:Blackwell Scientific. Swift, M.J. and P.A. Sanchez. 1984. Biological management of tropical soil fertility for sustained productivity. Nature and Resources 20 (4): 1-9. TropSoils. 1987. Annual report for 1985-86. Soil Science Department, North Carolina University. Vitousek, P.M. and R.L. Sanford, Jr. 1986. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol Syst. 17: 137-167. Wade, M.K. and P.A. Sanchez. 1983. Mulching and green manure applications for continuous crop production in the Amazon basin. Agron. J. 75: 39-45. Wiersum, K.F. 1984. Surface erosion under various tropical agroforestry systems. In C.L. O"Loughlin and A.J. Pearce (eds.), Symposium on effects of forest land use on erosion and slope-stability. Honolulu, Hawaii: East-West Center. Wiersum, K.F. 1985. Effects of various vegetation layers in an Acacia auriculiformis forest plantation on surface erosion in Java, Indonesia. In S.A. El-Swaify, W.C. Moldenhauer and A. Lo (eds.), Soil erosion and conservation. Ankeny, Iowa: Soil Conservation Society North America. Young, A. 1986a. Effects of trees on soils. In R.T. Prinsley and M.J. Swift (eds.), Amelioration of soils by trees: A review of current concepts and practices. London: Commonwealth Science Council. .1986b. The potential of agroforestry as a practical means of sustaining soil fertility. In R.T.Prinsley and M.J. Swift (eds.), Amelioration of soils by trees: A review of current concepts and practices. London: Commonwealth Science Council. .1986c. The potential of agroforestry for soil conservation. I. Erosion control (ICRAF Working Paper No. 42) II. Maintenance of fertility (ICRAF Working Paper 43. Nairobi: ICRAF. Young, A., R.J. Cheatle and P. Muraya. 1986. The potential of agroforestry for soil conservation. Part III. Soil Changes under Agroforestry (SCUAF): A predictive model. ICRAF Working Paper 44. Nairobi: ICRAF. Young, A. 1987. Soil productivity, soil conservation and land evaluation. Agroforestry Systems 5:277-292. |